1975年,在哈里·格伦德费斯特告诉我,需要以“每次一个细胞”的方式来研究大脑20年之后,我和同事们开始探索记忆的细胞学基础—一个人是如何能够终生都记得一次邂逅、一片美景、一场讲演或一份疾病诊断的呢?我们已经知道记忆源自神经环路中突触的变化:短时记忆来自功能性变化而长时记忆来自结构性变化。现在我们想钻入记忆谜团的更深处。我们想洞悉心理过程的分子生物学,想知道到底是哪些分子在短时记忆中起作用。带着这个问题,我们开始涉足一片完全未知的领地。
随着我对我们能够在海兔这一简单系统中找到记忆存储的分子基础的信心与日俱增,这段旅程就显得不那么令人生畏了。我们已经在海兔神经系统突触连接这个大迷宫中找到了一些门道,绘制了它的缩鳃反射的神经通路,证明了组成这一通路的突触能够被学习增强。毫无疑问,我们已经正确地走入了这个科学迷宫的外圈。现在我们想确定的是,短时记忆伴随的突触变化到底发生在这条神经通路的什么位置。
我们将注意力集中于传递触碰蜗牛虹吸管信息的感觉神经元与引发鳃收缩的运动神经元之间的关键突触上。现在我们想知道的是,构成这一突触的两个神经元如何导致了突触强度的变化。是感觉神经元对刺激的反应改变了,导致它的轴突终端释放的递质变得更多或更少了吗?还是运动神经元发生了改变,导致细胞中接收神经递质的受体数量增加,或者受体对递质的敏感性增加?
我们发现改变只是单方面的:在短时习惯化持续的几分钟里,感觉神经元释放出更少的神经递质,而在短时敏感化时,它释放出更多的神经递质。
后来我们知道,这种神经递质是谷氨酸,它是哺乳动物脑中的主要兴奋性递质。通过增加感觉细胞发送给运动细胞的谷氨酸数量,敏感化增强了运动细胞中激发的突触电位,这使得运动神经元更容易激发动作电位并导致鳃的收缩。 :::info 所以说速读,速看 能够短时释放更多神经递质,记忆印象更为深刻咯? ::: 感觉与运动神经元之间的突触电位只持续数毫秒,但我们观察到对海兔尾部的电击导致的谷氨酸释放和突触传递增加持续了很多分钟。这是如何发生的?
当我和同事们专注于解答这一问题时,我们注意到了一些有趣的现象。在感觉与运动神经元之间突触连接增强的同时,感觉细胞中存在着一个非常缓慢的突触电位,它持续数分钟,而不像运动神经元中典型的突触电位那样只存在数毫秒①。很快我们发现对海兔尾部的电击激活了第二类感觉神经元,它们接收来自尾部的信息。这些尾部感觉神经元激活了一组作用于虹吸管感觉神经元的中间神经元。是这些中间神经元引发了非常缓慢的突触电位。接着我们问自己:
这些中间神经元释放了什么神经递质?
这第二种神经递质如何导致了感觉神经元终端释放更多的谷氨酸,进而形成短时记忆的存储?
我们发现电击海兔尾部激活的中间神经元释放的是一种称作血清素的神经递质。这些中间神经元不仅与感觉神经元的胞体形成突触,还与突触前终端形成突触,因此它们不仅引发了一个慢突触电位,还增强了感觉细胞对运动细胞的谷氨酸释放。事实上,我们能够通过简单地施加血清素到感觉与运动神经元之间的连接,来模拟这一慢突触电位、突触强度的增强以及缩鳃反射的增强。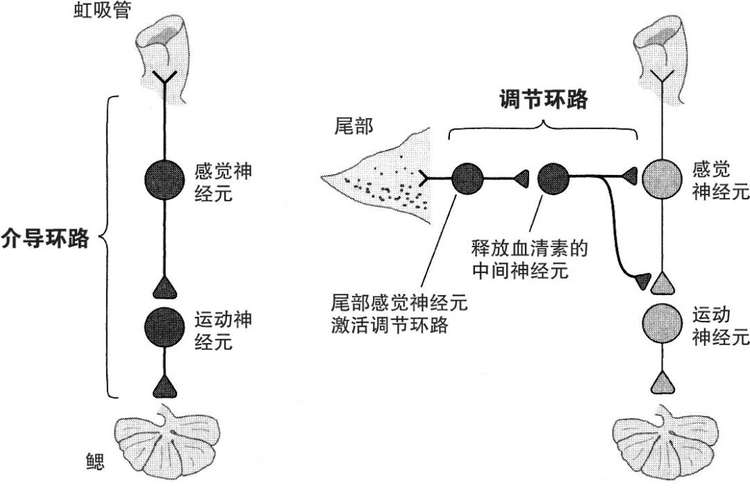
图16-1 脑中的两类环路。介导环路引发行为。调节环路作用于介导环路,调整它们的突触连接的强度。
我们把这些释放血清素的中间神经元称作调节性中间神经元,因为它们并不直接介导行为,而是通过增强感觉与运动神经元之间的连接强度来修饰缩鳃反射的强度。
这些发现让我们认识到,在行为与学习中存在两类重要的神经环路:我们之前描述过的介导环路和我们才刚开始深入研究的调节环路(图16-1)。
介导环路直接引发行为,因而属于康德式的,这些神经构造构成了行为中由基因和发育决定的那部分成分。介导环路由支配虹吸管的感觉神经元、中间神经元和控制缩鳃反射的运动神经元组成。在学习中,介导环路扮演学生的角色并获取新知识。
调节环路则属于洛克式,它充当老师。它并不直接引发行为,而是通过异突触调节感觉与运动神经元之间突触连接的强度,来对响应学习的行为进行微调。通过电击尾部这一与虹吸管截然不同的身体部位,调节环路得到激活,它教会海兔去注意对其安全很重要的虹吸管上的刺激。因此这个环路在本质上负责海兔的警觉或对显著性做出反应,正如我们之后会看到的,在更复杂动物身上的类似调节环路是记忆的必要成分。
血清素是敏感化的调质,这一事实让我感到很惊讶。1956年,我和多姆·普尔普拉做的第一个实验关注的就是血清素的作用。事实上在1956年春天,纽大医学院学生节那天,我还做过题为《传入皮层通路的血清素和 LSD 交互作用的电生理模式》的简短报告。排练时,好心的吉米·施瓦茨还帮助过我,给了我一些改进意见。我开始感慨人生就像一个圆圈。我已经几乎20年没研究过血清素了,而现在又带着新的眼光和热情重新回到了这里。 :::info 抗抑郁的药物,就有增加血清素的原理 ::: 一旦我们知道了血清素作为一种调节性递质,能够用来增强感觉神经元突触前终端的谷氨酸释放这一事实,下一步就是对**记忆存储**进行生化分析了。幸运的是,在这次科学旅程中,有吉米·施瓦茨这位杰出的向导与我同行。
回到纽大前,吉米在洛克菲勒大学研究过大肠杆菌,这是一种单细胞生物,许多现代生物化学和分子生物学的基本原理最初都源自对它的研究。1966年,他的兴趣转向了海兔,开始研究腹神经节的神经元使用的一种化学递质。1971年,我们开始一道研究起学习过程中伴随的分子作用。
在我接受生物学教育的第二个主要阶段中,吉米提供了不可估量的帮助。我们受到路易斯·弗莱克斯纳工作的影响,他几年前的研究表明小鼠和大鼠的长时记忆需要合成新蛋白质,短时记忆则不需要。蛋白质是细胞里的耕牛。它们组成了酶、离子通道、受体和运输机器。正如我们已经发现的,由于长时记忆涉及新连接的生长,因此生长需要新蛋白质成分的合成一点也不奇怪。 :::info 长时记忆需要生成新连接,需要蛋白质,所以学习,是一个不断新生的过程。 ::: 吉米和我开始在海兔身上测试这一想法,我们对虹吸管感觉细胞以及支配鳃的运动神经元的突触进行研究。如果突触变化平行于记忆变化,那么短时突触变化应该不需要合成新蛋白质。这与我们观察到的现象也相符。那么,是什么介导了这一短时变化呢?
卡哈尔已经提出,大脑是一个由特定通路上相互连接的神经元组成的器官。我在海兔身上介导反射行为的简单神经环路中也看到了这种值得注意的连接特异性。但吉米指出,这种特异性也适用于分子—通过原子结合而成的细胞功能的基本单元。生物化学家已经发现,一个细胞内的分子之间存在交互作用,而且这些化学反应是以特定顺序组织的,称作生化信号传导通路。这条通路以分子形式从细胞表面向细胞内部传递信息,很像神经细胞之间的信息传递。此外,这条通路还是“无线的”。在细胞内流动的分子与特异性分子伴侣识别并绑定,还调控后者的活性。
我和同事们不仅已经实现了我早期想在可能是最小的神经元群里捕捉到一个习得性反应的雄心壮志,而且我们已经在单个感觉细胞里捕捉到了记忆的一种简单形式的成分。但即使是单个海兔神经元也包括上千种不同的蛋白质及其他分子。这些分子里的哪个对短时记忆负责呢?
随着吉米和我开始探讨其中的可能性,我们注意到了一个想法,即响应尾部电击而释放的血清素可能通过在感觉细胞中发起特定序列的生化反应来增强感觉神经元谷氨酸的释放。
吉米和我要找的生化反应的序列必须服务于两个基本目的。第一,它们必须把血清素的短暂作用转化给感觉神经元中那些信号能够持续数分钟的分子。第二,这些分子必须从血清素发挥作用的细胞膜发出信号到感觉细胞的内部,特别是到涉及谷氨酸释放的轴突终端的特定区域。我们在1971年发表于《神经生理学期刊》的论文里详细描述了这些想法,并推测一个称作环腺苷酸的特定分子具备这种可能性。
什么是环腺苷酸?我们如何突发奇想将它列为可能的候选者?之所以想到环腺苷酸是因为研究者已经发现这个小分子是肌肉和脂肪细胞中信号传导的主要调控因子。吉米和我知道大自然是保守的—因此,适用于一种组织细胞中的机制很可能也适用于另一种组织细胞。克利夫兰市凯斯西储大学的厄尔·萨瑟兰德已经发现肾上腺素会引发脂肪和肌肉细胞膜表面的短暂生化变化,导致细胞内发生更持久的变化③。这一持续变化是由细胞内环腺苷酸数量的增强引起的。
萨瑟兰德的革命性发现称作第二信使信号传导理论。这一生化信号传导理论的核心是他在对激素做出反应的脂肪和肌肉细胞的表面发现了一类新的受体。在他之前,伯纳德·卡茨已经发现的神经递质门控受体属于离子通道型受体,这些受体在结合神经递质时,开启或关闭受体自带的离子通道的阀门,将化学信号转换为电信号。但称作代谢型受体的这一类新受体,自身没有可以开启或关闭的离子通道。取而代之的机制是,这些受体的一个区域从细胞膜外表面突出以识别来自其他细胞的信号,同时另一个区域从细胞膜内表面突出并吸引一个酶。当这些受体与细胞外的一个化学信使识别并结合时,它们会激活细胞内的腺苷酸环化酶,进而产生环腺苷酸。
这一过程的优点是可以极大地放大细胞的反应。当化学信使的一个分子与代谢型受体结合时,该受体刺激腺苷酸环化酶产生1000个环腺苷酸分子。接着环腺苷酸与一个关键蛋白质结合,触发整个细胞内一系列的分子反应。最终,腺苷酸环化酶继续产生环腺苷酸达数分钟的时间。因此,代谢型受体的作用比起离子通道型受体要更强大、更广泛也更持久。离子通道型作用通常只持续数毫秒,而代谢型作用能够持续几秒到几分钟—是前者的1000倍到10000倍。
为了区分代谢型受体空间上有差别的两种功能,萨瑟兰德将与代谢型受体细胞外部分结合的化学信使称作第一信使,而在细胞内受到激活并发出信号的环腺苷酸则是第二信使。他认为第二信使将来自细胞表面第一信使的信号传递到细胞内,并启动了整个细胞的反应(图16-2)。第二信使信号传导让我们想到,代谢型受体和环腺苷酸可能是把感觉神经元内慢突触电位与谷氨酸释放的增强以及最终短时记忆的形成连接到一起的神秘中介。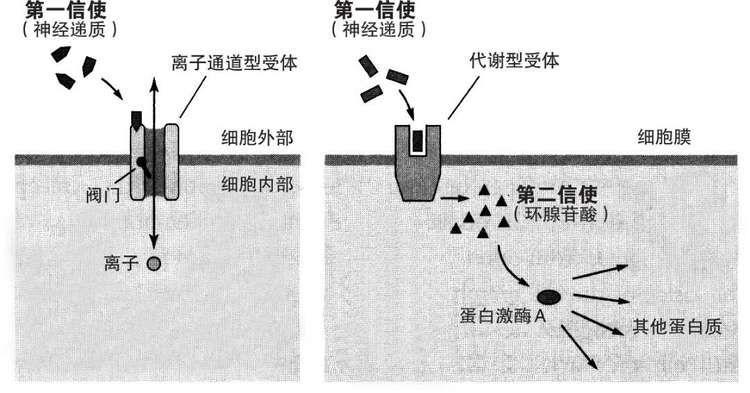
图16-2 萨瑟兰德的两类受体。离子通道型受体(左图)产生持续数秒的变化。代谢型受体(如血清素受体)通过第二信使发挥作用(右图)。它们在整个细胞内产生持续几秒到几分钟的改变。
1968年,华盛顿大学的爱德·克雷布斯提供了关于环腺苷酸如何产生广泛效应的最初洞见④。环腺苷酸结合并激活的酶被克雷布斯称作环腺苷酸依赖性蛋白激酶,或蛋白激酶 A(因为它是科学家发现的第一个蛋白激酶)。激酶通过给蛋白质添加一个磷酸分子来修饰它们,这一过程称作磷酸化。磷酸化激活一些蛋白质,同时让另外一些蛋白质失活。克雷布斯发现磷酸化很容易逆转,因此能够作为一个简单的分子开关,打开或关闭一个蛋白质的生化活性。
克雷布斯接下来研究了这个分子开关是如何工作的。他发现蛋白激酶 A 是一个由4个单元组成的复杂分子,包括两个调控单元和两个催化单元。催化单元用于实现磷酸化,而调控单元通常“坐”在它们上面以抑制它们。调控单元包含结合环腺苷酸的位点。当环腺苷酸在细胞内的浓度上升时,调控单元结合这些过量的分子。这一作用改变了它们的构型并使得它们脱离催化单元,让催化单元自由地去将靶蛋白磷酸化。
这些发现让我们意识到一个关键问题:萨瑟兰德和克雷布斯发现的这一机制是特异性地只对脂肪和肌肉细胞的激素起作用,还是也对包括脑中含有的递质在内的其他递质也起作用?如果是后者的话,这将代表突触传递的一个前所未知的机制。 :::info 可迁移性 ::: 这里我们得到了保罗·格林加德工作的帮助。他是一位受过生理学训练的天才生化学家,当时刚从他担任生物化学主任的嘉基制药研究实验室调到耶鲁大学不久。在去耶鲁之前,他在萨瑟兰德任教的系里待了一年。他认识到脑中一个潜在的新信号传导机制的重要性,于1970年开始研究大鼠脑中的代谢型受体。现在出现了一个奇妙的巧合,将阿尔维德·卡尔森、保罗·格林加德和我在科学之旅上连到一起,并让我们仨在2000年到达斯德哥尔摩,凭借神经系统信号转换方面的贡献分享了诺贝尔生理学或医学奖。
1958年,伟大的瑞典药理学家阿尔维德·卡尔森发现了多巴胺是神经系统的一种递质。接着他进一步发现,当兔子体内的多巴胺浓度降低时,它会出现类似帕金森氏病的症状。当格林加德开始探索脑中的代谢型受体时,他从一个多巴胺的受体入手,发现它刺激的一个酶增加了脑中的环腺苷酸并激活了蛋白激酶 A!
基于这些指引,吉米·施瓦茨和我发现环腺苷酸第二信使信号传导同样是在敏感化时由血清素开启的。我们已经知道,电击海兔尾部会激活调节性中间神经元释放血清素。接着,血清素将会增加感觉神经元突触前终端中环腺苷酸的产生并持续几分钟(图16-3)。于是,这些事情都联系到了一起:环腺苷酸的增加持续的时间和慢突触电位持续的时间、感觉与运动神经元直接的突触电位的增加持续的时间和动物对电击做出的加剧行为反应持续的时间都是一样长的。
图16-3 短时记忆的生化步骤。电击海兔尾部会激活中间神经元释放化学信使血清素到突触。在穿过突触间隙后,血清素与感觉神经元的受体结合,导致环腺苷酸的产生(步骤1)。环腺苷酸让蛋白激酶 A 的催化单元得到自由(步骤2)。蛋白激酶 A 的催化单元增强了神经递质谷氨酸的释放(步骤3)。
环腺苷酸参与形成短时记忆的第一个直接证据出现在1976年,当意大利籍博士后研究员马尔塞洛·布鲁内利加入我们实验室的时候。布鲁内利检验了一个想法,即当血清素发送信号给感觉神经元增加环腺苷酸的浓度时,该细胞提升了从其终端释放谷氨酸的量。我们直接注射环腺苷酸到海兔的感觉细胞,发现谷氨酸的释放量出现了大幅度增加,于是,感觉细胞与运动神经元之间的突触增强了。事实上,注射环腺苷酸完美地模拟了通过给感觉神经元施加血清素或电击海兔尾部所引发的突触强度的增加。这一令人瞩目的实验不仅将环腺苷酸与短时记忆联系了起来,而且给我们提供了关于学习的分子机制的第一手洞见。从捕捉短时记忆的基本分子成分起步,现在我们已经能够利用它们来模拟记忆的形成了。
1978年,吉米和我开始与格林加德合作。我们仨想知道环腺苷酸是不是通过蛋白激酶 A 来对短时记忆产生效应的。我们将这个蛋白质拆开,并只把通常会造成磷酸化的催化单元直接注入感觉神经元。我们发现,这个单元完成的正是环腺苷酸的功能—它通过促进谷氨酸的释放而增强了突触连接。接着,为了确定我们的方向是正确的,我们将蛋白激酶 A 的一种抑制剂注入了感觉神经元,发现它确实阻断了血清素促进谷氨酸释放的能力。发现环腺苷酸和蛋白激酶 A 是增强感觉与运动神经元之间连接的充分必要条件后,我们就能够确定导致短时记忆存储的第一条生化反应链了(图16-4)。
图16-4 参与短时记忆的分子。施加血清素到感觉神经元的终端(1)、注射环腺苷酸到神经元内(2)以及注入蛋白激酶 A 的催化单元(3)都导致了神经递质谷氨酸释放的增加。这表明上述三种物质分别参与了形成短时记忆的通路。
然而,这并没有告诉我们血清素和环腺苷酸如何引发慢突触电位,或这一突触电位如何与促进谷氨酸的释放相关。1980年,我在巴黎遇见了史蒂文·西格尔鲍姆,当时我正在法兰西公学院开展一系列研讨会。史蒂夫⑤是一个具有技术天赋的年轻生物物理学家,专长是研究单个离子通道的性质。我们简直是一拍即合,而且或许是命中注定,他刚刚接受了哥大药理学系的一个职位。于是我们决定待他来到纽约后开始合作探索慢突触电位的生物物理学性质。
史蒂夫发现了环腺苷酸和蛋白激酶 A 的一个靶点:对血清素做出反应的感觉神经元上的一个钾离子通道。我们把它称作 S 通道,因为它对血清素做出反应,而且它是史蒂夫·西格尔鲍姆发现的⑥。当神经元处于静息态时通道是开放的,并对静息态时的膜电位有贡献。史蒂夫发现这个通道出现在突触前终端,他能够通过向细胞膜外施加血清素(第一信使)或者向细胞内部施加环腺苷酸(第二信使)或蛋白激酶 A 来让通道关闭。关闭钾离子通道会引发慢突触电位,这又把我们的注意力首先放到了环腺苷酸上。
关闭这一通道还有助于谷氨酸的释放。当这个通道开放时,它和其他钾离子通道一起对静息态膜电位以及在动作电位的下行阶段对钾离子的外流做出贡献。但当它被血清素关闭时,离子外流变慢,通过减缓下行阶段而稍微增加了动作电位的持续时间。史蒂夫发现动作电位的延长使得钙离子有更多时间流向突触前终端—而正如卡茨在乌贼的巨大突触中显示过的,钙离子对谷氨酸的释放很关键。此外,环腺苷酸和蛋白激酶 A 直接作用于释放突触囊泡的装置,因此进一步刺激了谷氨酸的释放。
这些令人兴奋的有关环腺苷酸的研究结果很快得到了来自果蝇学习的重要遗传学研究的补充,这是一项在之前的半个多世纪中一直很热门的研究。1907年,哥大的托马斯·亨特·摩尔根⑦开始使用果蝇作为遗传学研究的模式生物,因为它的体型小而且繁殖周期短(12天)。这被证明是一个明智的选择,因为果蝇只有4对染色体(相比之下人类有23对),使得它成为一种相对容易用于遗传学研究的动物。一直以来显而易见的是,动物的许多物理特征—体型、眼睛的颜色以及速度等—都是遗传的。如果外在的物理特征能够遗传,那么大脑产生的心理特征是否也能够遗传呢?基因是否在心理过程—比如记忆—中扮演角色?
加州理工学院的西摩·本泽是运用现代技术研究这个问题的第一人。1967年,他开启了一系列绝妙的实验,用化学物质处理果蝇以造成单个基因随机的突变。接着他检查这些突变对学习与记忆的影响。为了研究果蝇的记忆,本泽的学生奇普·奎因和亚丁·杜戴采用了经典条件作用程式。他们将果蝇置于一间小室内,让它们先后暴露在两种气味中。然后果蝇在第一种气味出现时受到电击,它们学会了避开这种气味。接着,果蝇被置于另一间小室,小室两头分别放有一种气味源。被条件化的果蝇避开了包含第一种气味的一头而飞向包含第二种气味的一头。
奎因和杜戴利用这一训练程式来鉴别出一类果蝇,它们缺乏记住伴随电击的第一种气味的能力。到1974年,他们筛选了上千种果蝇,并找到了存在短时记忆缺陷的第一个突变体。本泽把这个突变体称作“傻瓜”。1981年,本泽的学生邓肯·拜尔斯跟进了对海兔的研究,开始考察“傻瓜”的环腺苷酸通路并发现一个基因上的突变对处理环腺苷酸负责。发生这种突变将造成果蝇体内积累过多的环腺苷酸,它的轴突很可能已经饱和,使得它们对进一步的改变不敏感并阻碍其发挥最佳功能。接下来其他一些有关记忆的基因突变也得到了鉴定。它们同样参与了环腺苷酸通路。
海兔和果蝇研究得到的互相印证的结果—两种截然不同的实验动物,通过不同的方法测试了不同类型的学习—非常令人安心。合到一起,它们清楚地表明,简单形式的内隐记忆的细胞机制很可能在许多物种(包括人类)的许多不同形式的学习中都是相同的,因为这些机制在进化过程中是保守的。生物化学和之后的分子生物学会是揭示不同有机体的生物构造共同特征的强大工具。
海兔和果蝇研究中的发现还加强了一个重要的生物学原理:进化不需要新的特异性分子来产生一个新的适应性机制。环腺苷酸通路并不是记忆存储独有的。正如萨瑟兰德所展示的那样,它甚至不是神经元独有的:肠道、肾脏和肝脏都使用环腺苷酸通路来产生持久的代谢变化。事实上,在已知的全部第二信使中,环腺苷酸系统可能是最原始的一种。它也是最重要的,在某些情况下,如在单细胞生物中,它甚至是唯一的第二信使系统,比如在大肠杆菌中它发送饥饿信号。因此,构成记忆基础的生化反应并非专门用来支持记忆的。神经元仅仅是招募了一个在其他细胞中用于其他目的的有效信号传导系统,并用它来产生记忆存储所需的突触强度的变化。
正如分子遗传学家弗朗索瓦·雅各布曾指出的,进化并不是一个原创设计师,它不会发明一套全新的方案来解决新问题。进化是一个修补匠。它反复地使用相同的基因,只是每次的用途都略有不同。进化通过改变现有条件并筛选出基因结构中的随机突变来发挥作用,被筛选的这些突变会使一个蛋白质变得稍有不同或者改变该蛋白质在细胞中配置的方式。大多数突变是中性的甚至有害的,经不起时间的考验而消失了。只有少量能增加个体存活率和繁殖能力的突变才有可能被保留下来。雅各布这样写道:
大家经常把自然选择比作工程师。然而这一比较似乎并不合适。首先……工程师是根据事先制订好的计划来工作的。其次,一个工程师在组装一个新结构时并不必然是在旧结构的基础上进行的。电灯不是源自蜡烛,喷射发动机也不是从内燃发动机发展而来。……最后,一个工程师,至少一个优秀的工程师,是从头开始制作一个物件,并在当时技术允许的条件下尽可能地让这个物件达致完美。
与工程师不同,进化并不是从头开始进行创造。它在现有的基础上工作,有时调整一个原有的系统让它具有新功能,有时将几个系统合并起来,形成一个更复杂的体系。不过如果谁一定要打比方的话,也许修补匠是个比工程师更好的选择,我们在法语里把这种修修补补的活动称作 bricolage。工程师的工作依赖于他的原材料和适合他的项目的工具,而修补匠打理的是边角料。……他会用上周遭所能找到的任何玩意,旧纸板、几根线、碎木料或金属片,来制造出某种可用之物。修补匠捡起恰好在他仓库里的一个物件,并赋予它意想不到的功用。给他一辆老汽车的方向盘,他会做出一台风扇;给他一张破桌子,他能做出一把遮阳伞。
在活着的有机体中,对现有的分子稍加修饰并调整它们与其他分子的交互作用,就可以实现新功能。因为人类的心理过程长期以来都被认为是独一无二的,一些早期的脑研究者期待在我们的灰质中找到许多潜藏的蛋白质新种类。然而令人惊讶的是,科学研究发现几乎没有哪种蛋白质是人脑独有的,即便有,它也不具有独特的信号传导系统。几乎脑中所有的蛋白质,都在身体的其他细胞中存在着执行相似目的的亲戚。这一点即便对执行大脑独有功能的蛋白质,比如作为神经递质受体的那些蛋白质也是成立的。所有的生命,包括构成我们思想和记忆的基质,都是由相同的构件组成。
我在出版于1976年的《行为的细胞基础》一书里首次总结出了关于短时记忆的细胞生物学的贯通性主张。在书中我清楚地说明了我的信念—甚至可以说是发表了我的宣言—要理解行为,就必须把已被证明在生物学其他领域非常有效的激进还原论取向同样运用到行为研究中。大约在同一时间,斯蒂夫·库夫勒和约翰·尼科尔斯出版了《从神经元到脑》,这本书强调了细胞取向的重要性。他们用细胞生物学来解释神经细胞如何工作以及它们如何在脑中形成环路,而我则用细胞生物学将脑与行为连接起来。斯蒂夫也察觉了这种连接的存在,并预见到神经生物学领域正要向前迈出一大步。
1980年8月,我和斯蒂夫被奥地利生理学会授予荣誉会员称号,一同受邀去维也纳访问,能和他同行让我感到非常高兴。斯蒂夫是于1938年逃离维也纳的。威廉·奥尔瓦尔德把我们介绍给维也纳大学医学院的全体教师。这位自命不凡的学者在科研上几无建树,对于我们两个维也纳之子被迫逃离这个国家的经历,他也表现得无动于衷。他高兴地提到库夫勒曾就读于这所医学院,而我曾住在位于这所大学拐角处的塞弗林巷,却对我们在这期间的实际经历避而不谈。他的这一态度伤害了斯蒂夫和我,我们没有对他的言论做出回应。
两天后,我们从维也纳乘船沿着多瑙河来到布达佩斯,参加生理学国际会议。这是斯蒂夫参加的最后一次重要会议,他在会上做了非常精彩的报告。不久后的1980年10月,他在一次长时间的游泳之后,突发心脏病,死在了位于马萨诸塞州伍兹霍尔的周末居所内⑧。
听到这个消息后,整个神经科学界都深感悲痛。我们中大多数人都受惠于他并在某些方面依赖于他。斯蒂夫的得意门生杰克·麦克马汉说出了我们许多人的心声:“他怎能就这样丢下我们不管了呢?”
我是那年的神经科学学会主席,与项目委员会一道负责组织11月的年会。斯蒂夫去世仅几个星期后,这次会议在洛杉矶开幕,有大约一万名神经科学家参加。戴维·休伯尔发表了非凡的悼词。他用幻灯片图文并茂地展示了斯蒂夫的预见性、洞察力和慷慨,以及他对我们所有人的意义。我认为从那以后,美国社会再也没有哪个人能像斯蒂夫·库夫勒这样有影响力并深受爱戴了。杰克·麦克马汉主编了一本纪念他的文集,我在其中写道:“写下这些文字时,我仍然深深地感觉到他的存在。继奥尔登·斯宾塞之后,还没有哪位同事的离开能让我感到如此悲伤和怀念。”
斯蒂夫·库夫勒的去世标志着一个时代的结束。在过去的这个时代里,神经科学界规模相对还比较小、科学家们所关注的还只是作为脑组织单元的脑细胞而已。斯蒂夫去世时恰逢分子生物学和神经科学融合的时代,这一融合极大地扩展了这一领域的范围和参与其中的科学家的数量。我本人的工作也反映了这一变化:我在1980年停止了大部分对学习与记忆的细胞和生化研究。那时我已经清楚地认识到,对单个学习试次做出反应的环腺苷酸的增加和血清素引发的递质释放的增加只持续几分钟。持续几天到几星期的长时程易化必须有别的什么东西参与,可能是基因表达的变化以及解剖学变化。因此我转向了基因研究。
我已经为走出这一步做好了准备。长时记忆开始点燃我的想象力。一个人怎么能够把小时候发生的事记一辈子?迪尼丝的母亲萨拉·贝斯特林,用她对装饰艺术—新艺术运动⑨的家具、花瓶和灯饰—的品位感染了迪尼丝和她的弟弟让–克劳德以及他们的配偶和孩子。虽然她极少跟我谈起我的科研,但她一定已经感觉到我准备去研究基因和长时记忆了。
在1979年11月7日我50岁生日那天,她送给我一个泰普利斯特制作的漂亮维也纳花瓶(图16-5),并附上了下面这张便笺:

若有收获,就点个赞吧
0 人点赞
