DNA的复制
- 复制(replication) 是以DNA为模板的DNA合成,是基因组的复制过程。在这个过程中,亲代DNA作为合成模板,按照碱基配对原则合成子代分子,其化学本质是酶促脱氧核苷酸聚合反应。
- DNA复制的主要特征
- 半保留复制 (semi-conservative replication)
- 双向复制 (bidirectional replication)
- 半不连续复制 (semi-discontinuous replication)
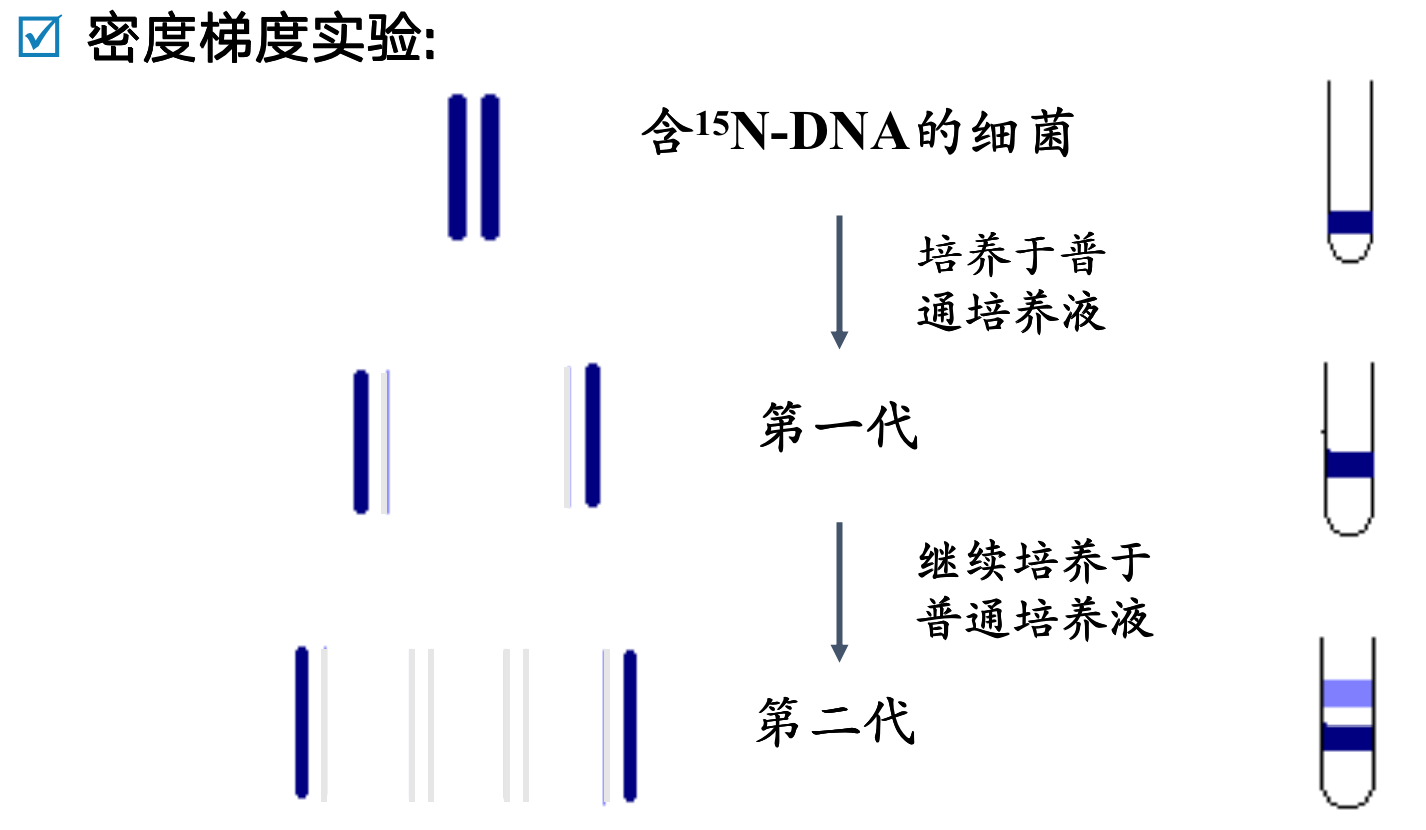
DNA以半保留方式进行复制
- 在复制时,亲代双链DNA解开为两股单链,各自作为模板,依据碱基配对规律,合成序列互补的子链DNA双链
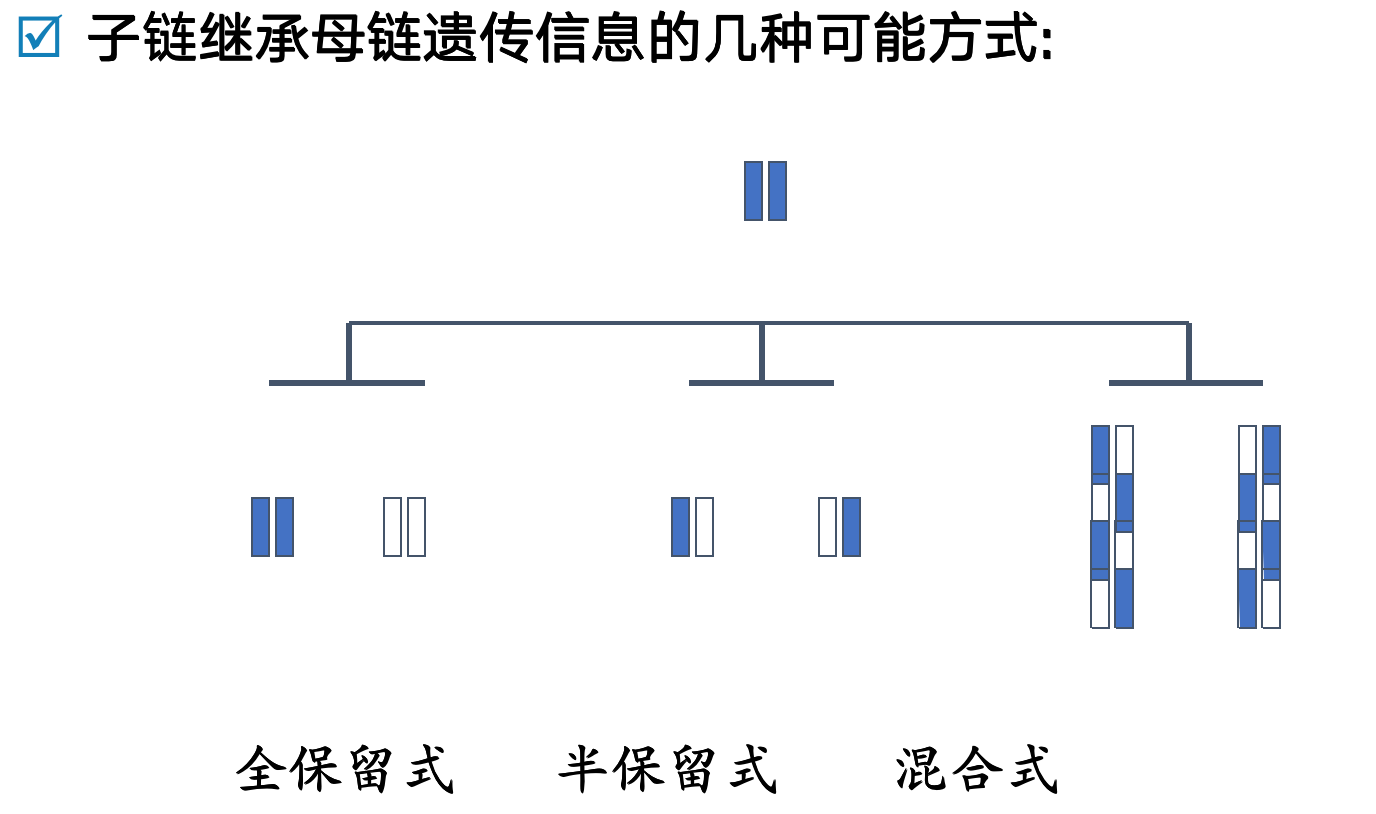
- 半保留复制的意义
- 依据半保留复制的方式,子代DNA中保留了亲代的全部遗传信息,亲代与子代DNA之间碱基序列的高度一致
- 遗传的保守性,是物种稳定性的分子基础,但不是绝对的
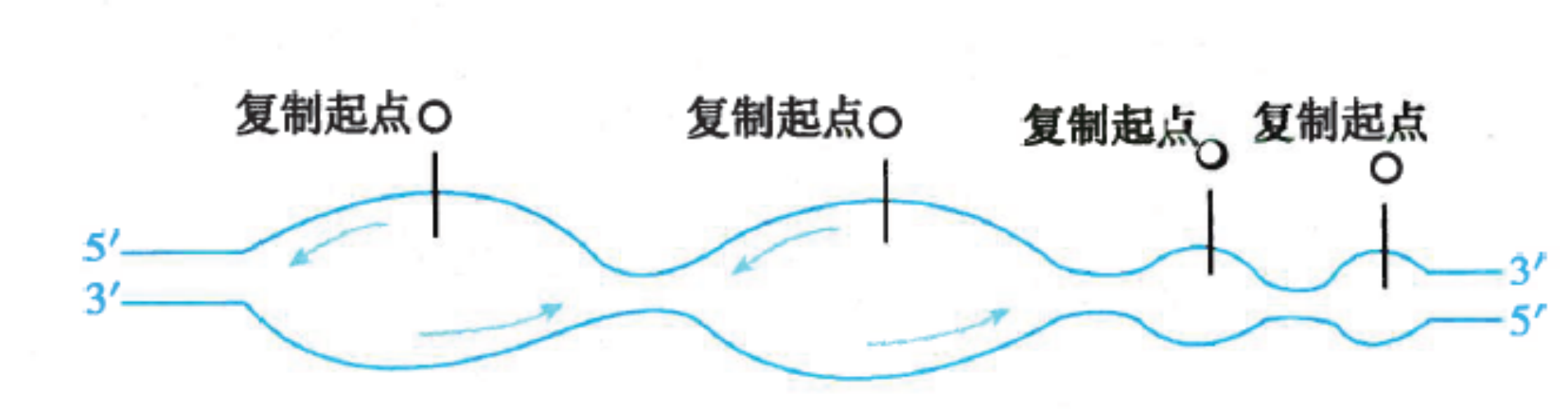
DNA复制从起始点双向进行
- 原核生物基因组是环状DNA,只有一个复制起点(origin)。复制从起点开始,向两个方向进行解链,进行的是单点起始双向复制
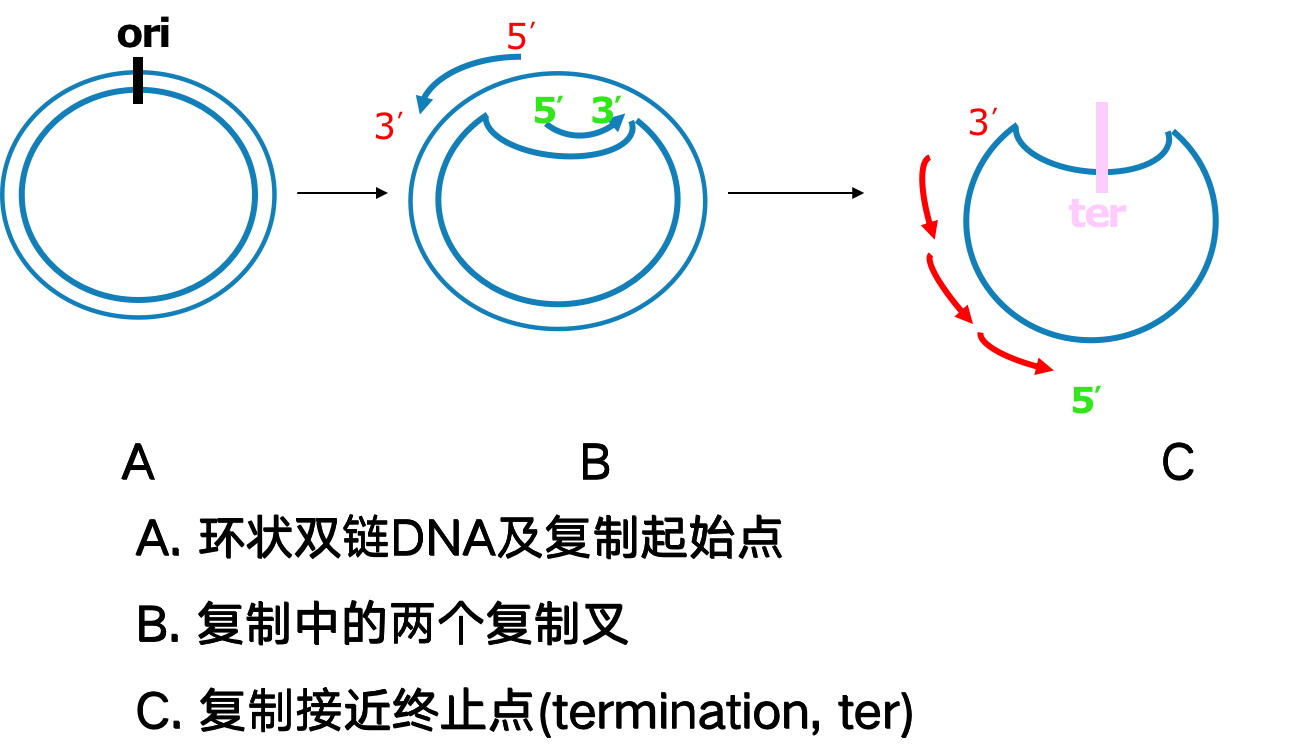
真核生物每个染色体又有多个起点,呈多起点双向复制特征。每个起点产生两个移动方向相反的复制叉,复制完成时,复制叉相遇并汇合连接。从一个DNA复制起点起始的DNA复制区域称为复制子(replicon)。复制子是含有一个复制起点的独立完成复制的功能单位
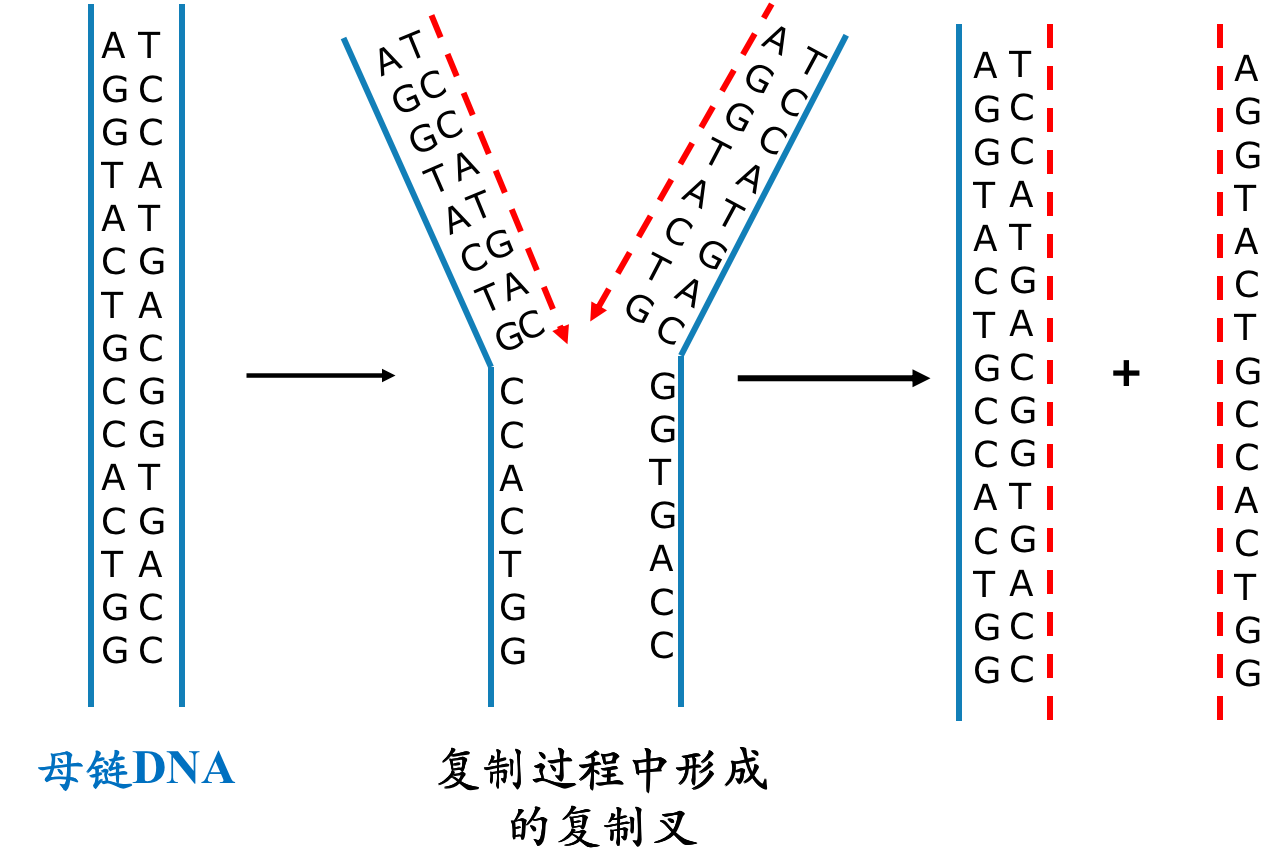
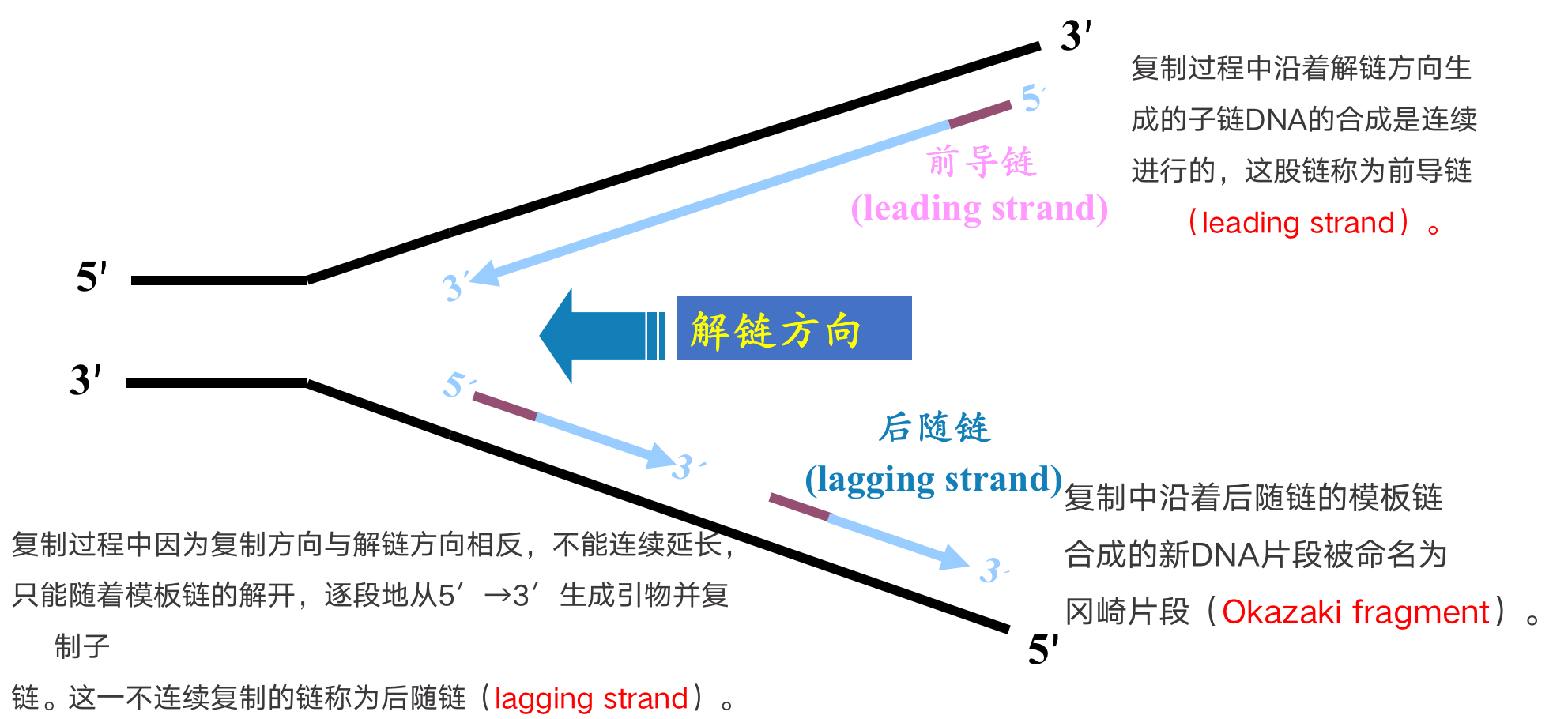
DNA复制以半不连续方式进行
DNA复制的酶学和拓扑学
参与DNA复制的物质
- 底物(substrate): dATP, dGTP, dCTP, dTTP
- 聚合酶(polymerase): 依赖DNA的DNA聚合酶,简写为 DNA-pol
- 模板(template): 解开成单链的DNA母链
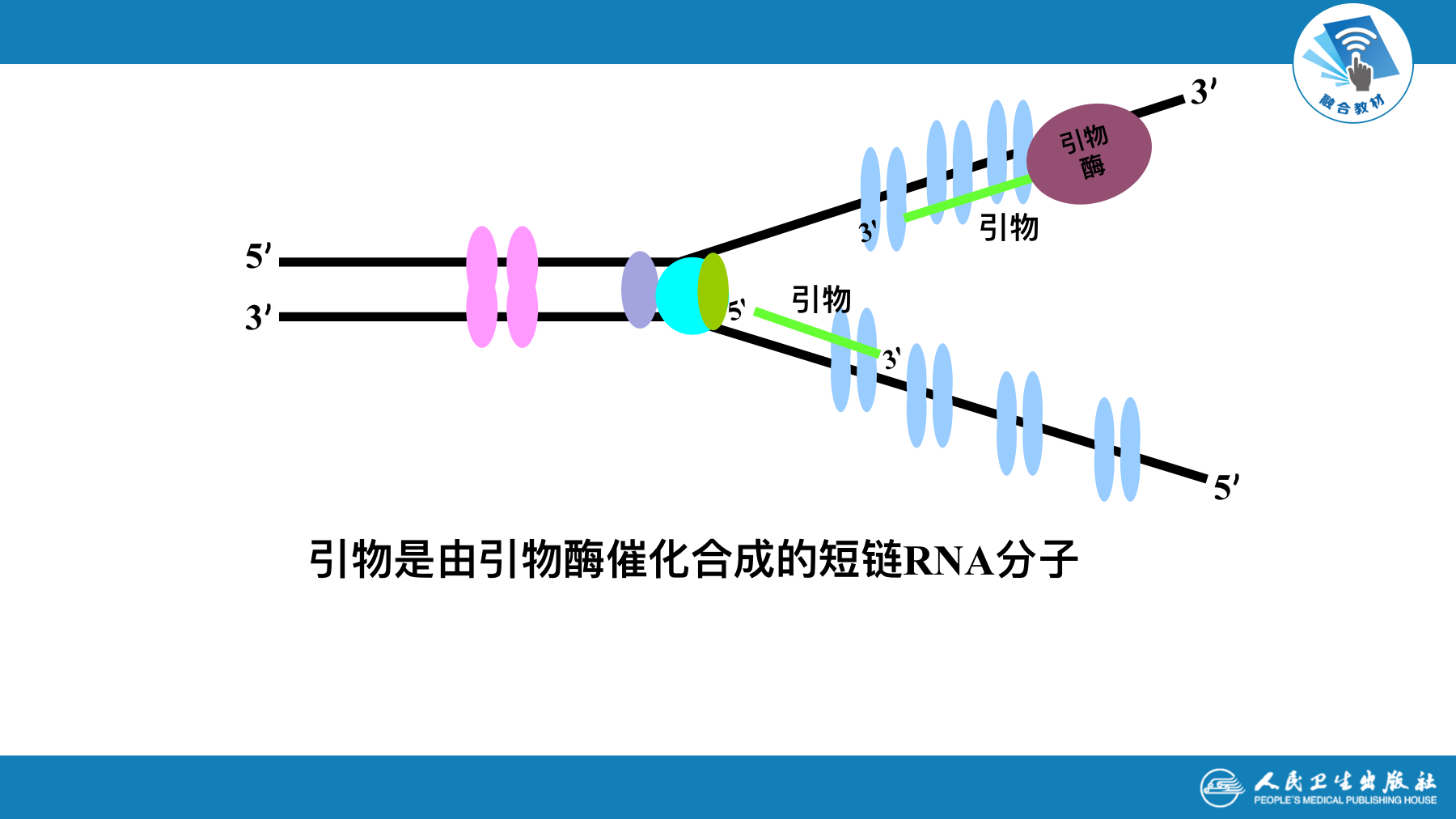
- 引物(primer): 提供3’-OH末端使dNTP可以依次聚合
- 引物酶:引物是引物酶催化合成的短链RNA分子
- 其他的酶和蛋白质因子
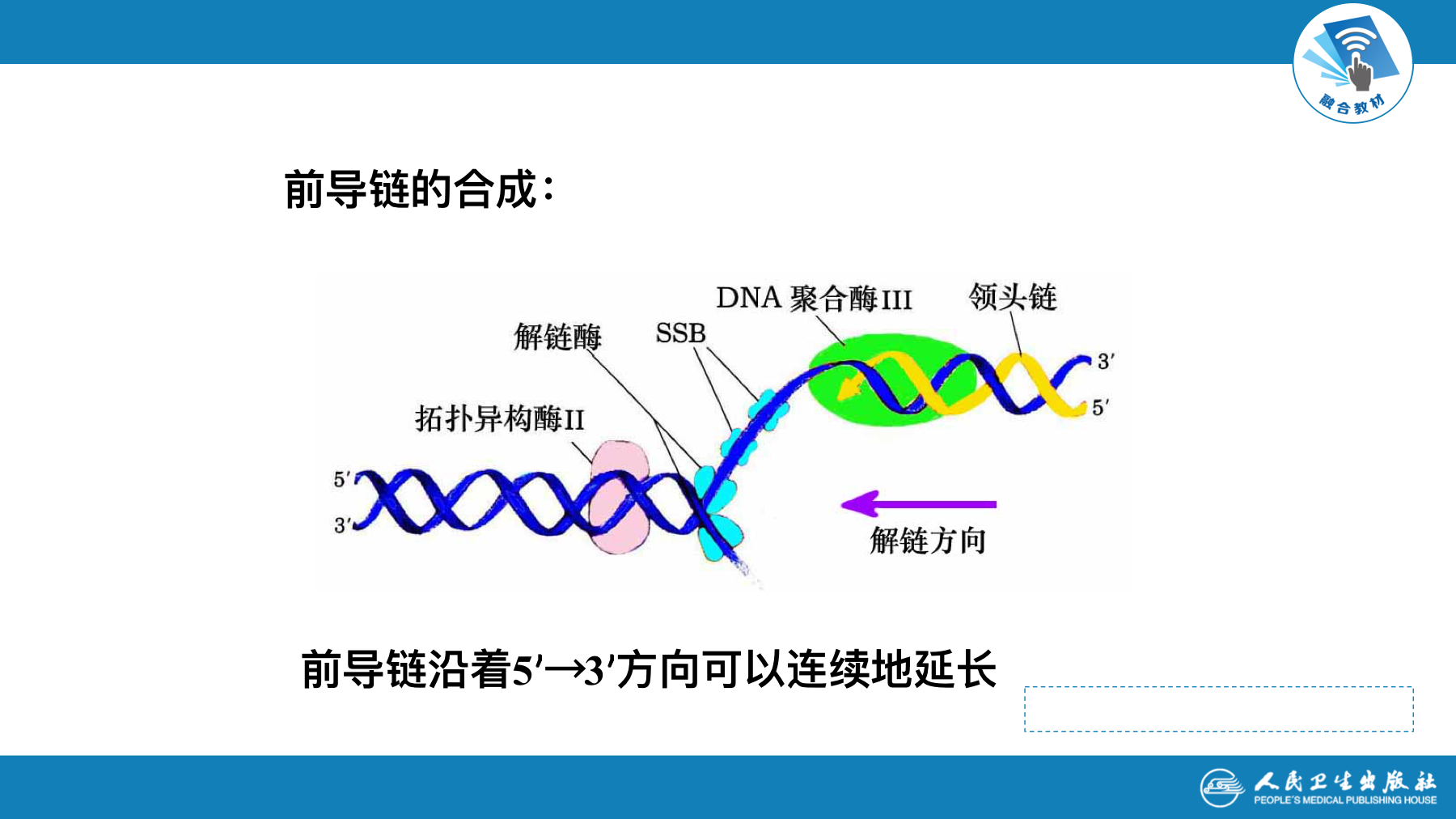
- 新链的延长只可沿5’-3’方向进行
- DNA新链生成需RNA引物和DNA聚合酶
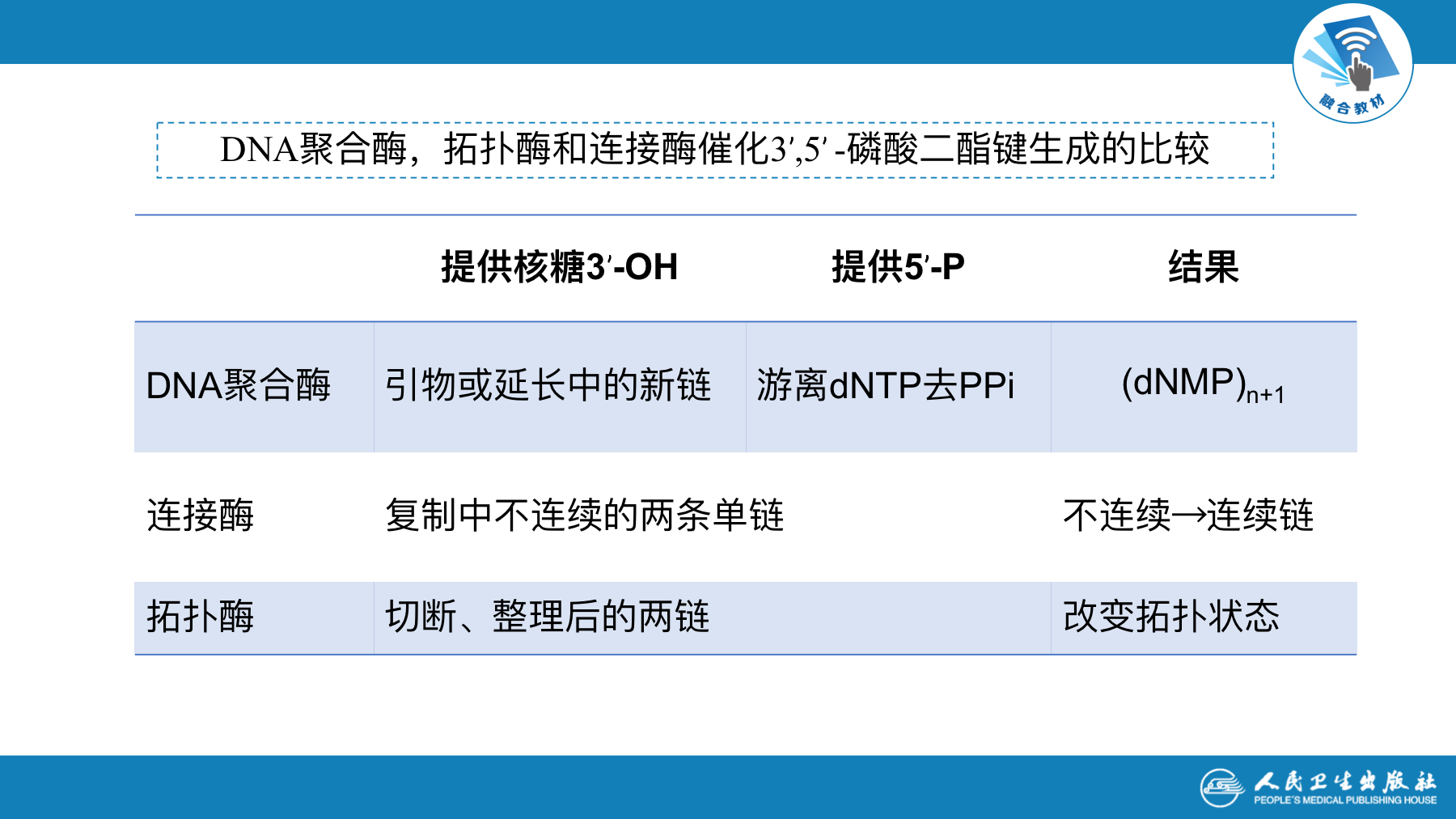
DNA聚合酶催化脱氧核苷酸间的聚合
- 全称:依赖DNA的DNA聚合酶 (DNA-dependent DNA polymerase)
- 简称:DNA-pol
- 功能:
- 聚合作用:在引物RNA-OH末端,以dNTP为底物,按模板DNA上的指令,即A与T,C与G的配对原则,逐步逐个、连续地将dNTP加到延伸中的DNA分子3-OH末端,逐步合成延长中的子链DNA。这是DNA聚合酶的主要作用
- 校对作用:从3’→5’方向识别和切除不配对的DNA生长链末端的核苷酸
- 切除修复作用:该活性是从5’→3’方向水解DNA延长链前方的DNA链(即只对DNA上双链处的磷酸二酯键有切割作用),主要产生5—脱氧核苷酸。在DNA损伤的修复中可能起着重要作用。
- 活性
- 5’-3’的聚合活性
- 核酸外切酶活性

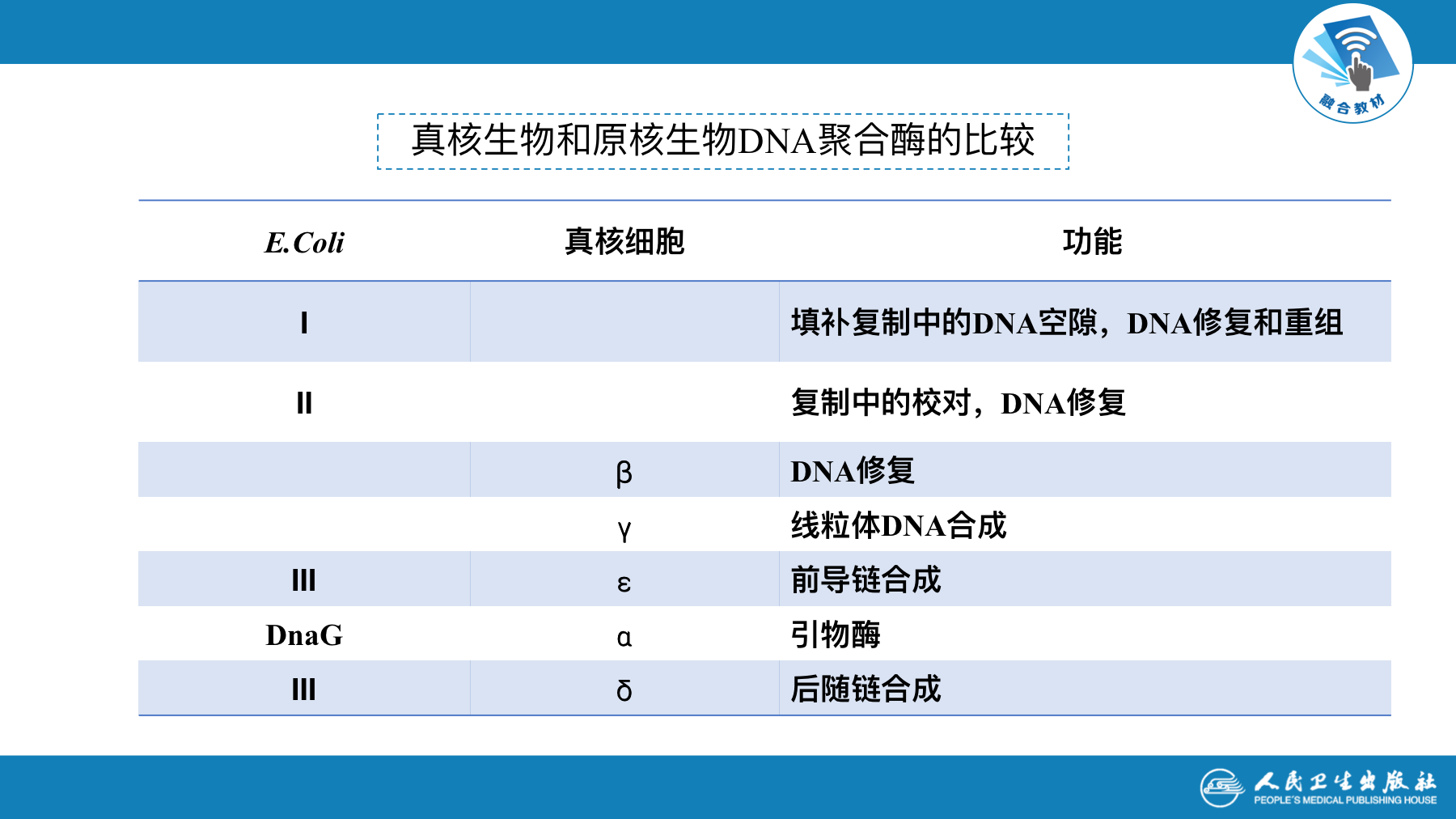
- 原核生物至少有5种DNA聚合酶
- DNA-polⅠ
- DNA-polⅡ
- DNA-pol Ⅲ
- DNA-pol Ⅳ
- DNA-pol Ⅴ
- 原核生物的DNA聚合酶 | | DNA-pol Ⅰ | DNA-pol Ⅱ | DNA-pol Ⅲ | | —- | —- | —- | —- | | 分子量(kD) | 109 | 120 | 250 | | 组成 | 单肽链 | ? | 多亚基不对称二聚体 | | 分子数/细胞 | 400 | ? | 20 | | 5’->3’ 核酸外切酶活性 | 有 | 无 | 无 | | 3’->5’ 核酸外切酶活性 | 有 | 有 | 无 | | 5’->3’聚合酶活性 | 有 | 无 | 有 | | 基因突变后的致死性 | 可能 | 不可能 | 可能 |
DNA聚合酶III全酶结构
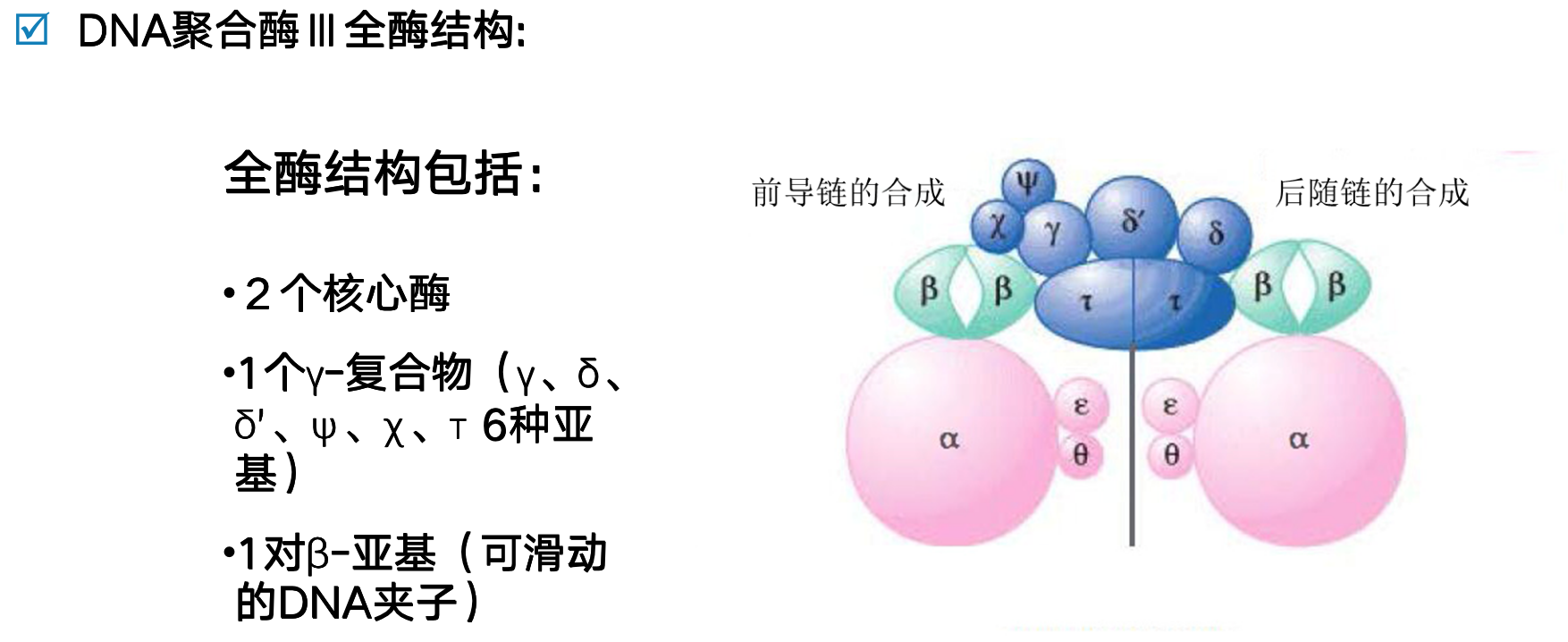
- 核心酶由∂、ß和θ亚基组成
- ∂亚基(130000)主要功能是合成DNA
- ß亚基具有3’-5’外切酶活性(复制保真性所必需)ß亚基可增强其活性
- θ亚基可能起组装作用
- 两侧的β亚基发挥夹稳DNA模板链,并使酶沿模板滑动的作用

DNA聚合酶I

DNA聚合酶II

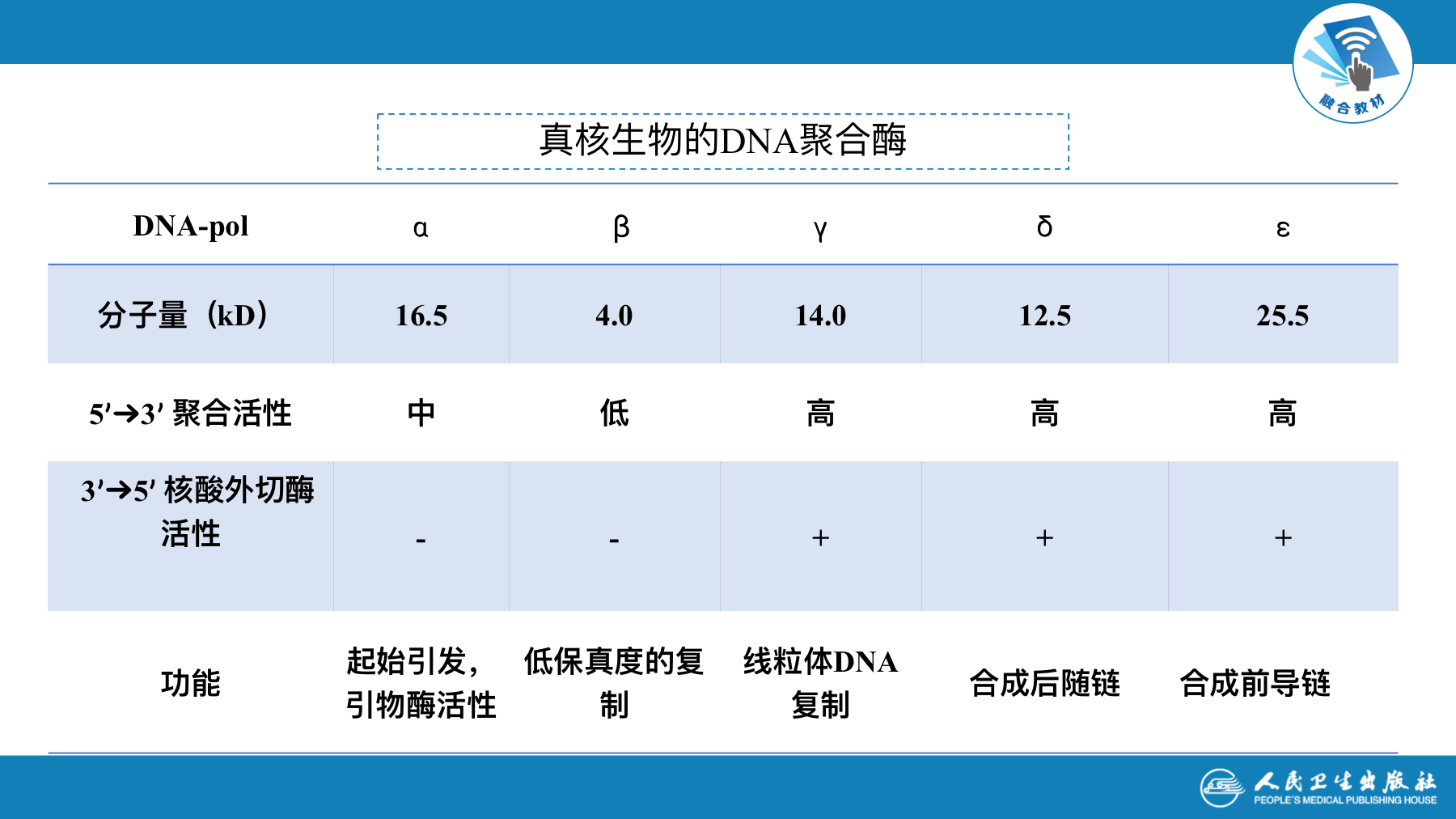
常见的真核细胞DNA聚合酶有5种
| DNA-pol ∂ | 起始引发,有引物酶活性 |
|---|---|
| DNA-pol ß | 参与低保真度的复制 |
| DNA-pol γ | 在线粒体DNA复制中起催化作用 |
| DNA-pol δ | 合成后随链 |
| DNA-pol ε | 合成前导链 |
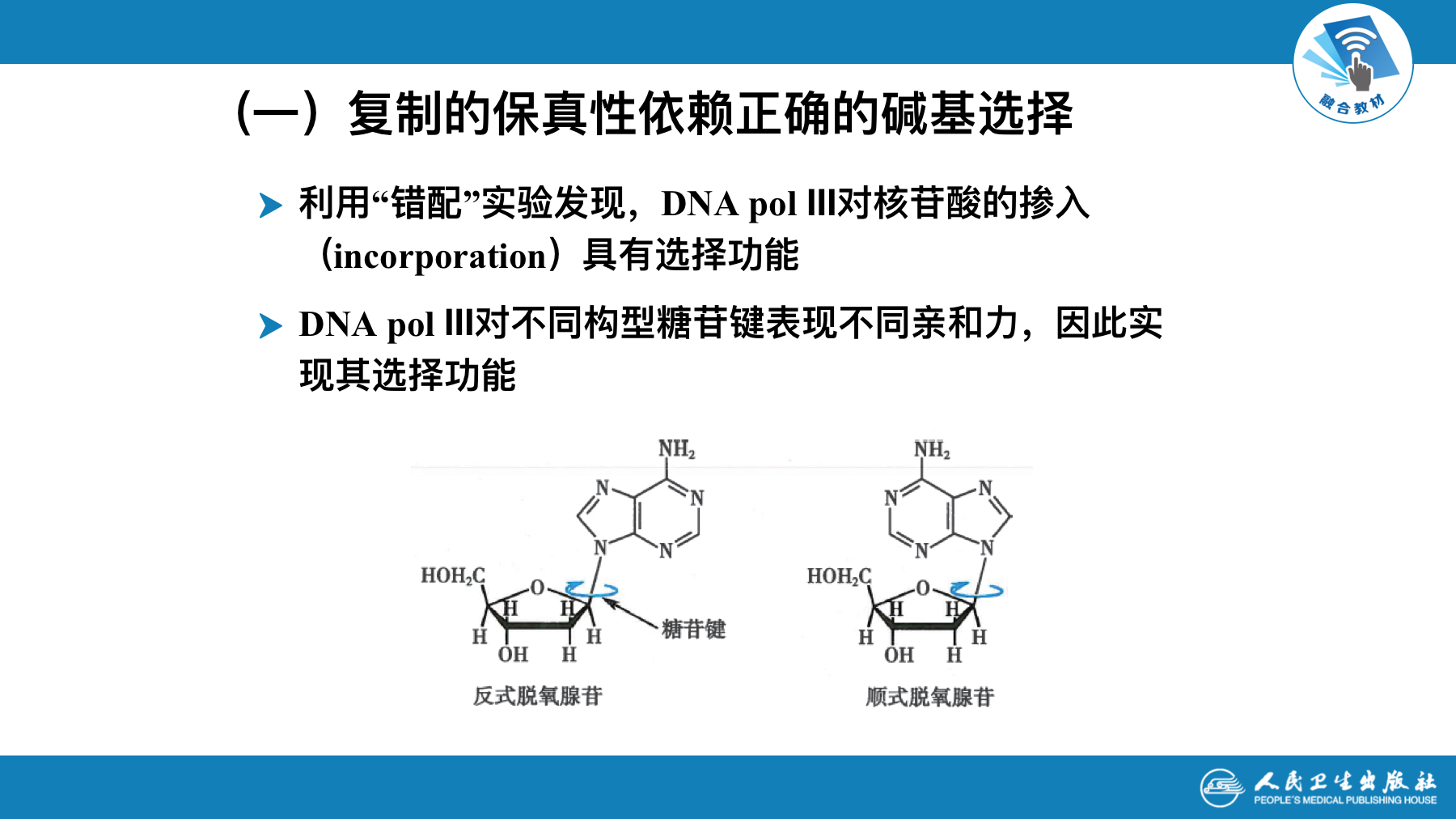
DNA聚合酶的碱基选择和校对功能
- DNA复制的保真性至少要依赖三种机制
- 遵守严格的碱基配对规律
- 聚合酶在复制延长时对碱基的选择功能
- 复制出错时有即时校对功能
复制的保真性依赖正确的碱基选择
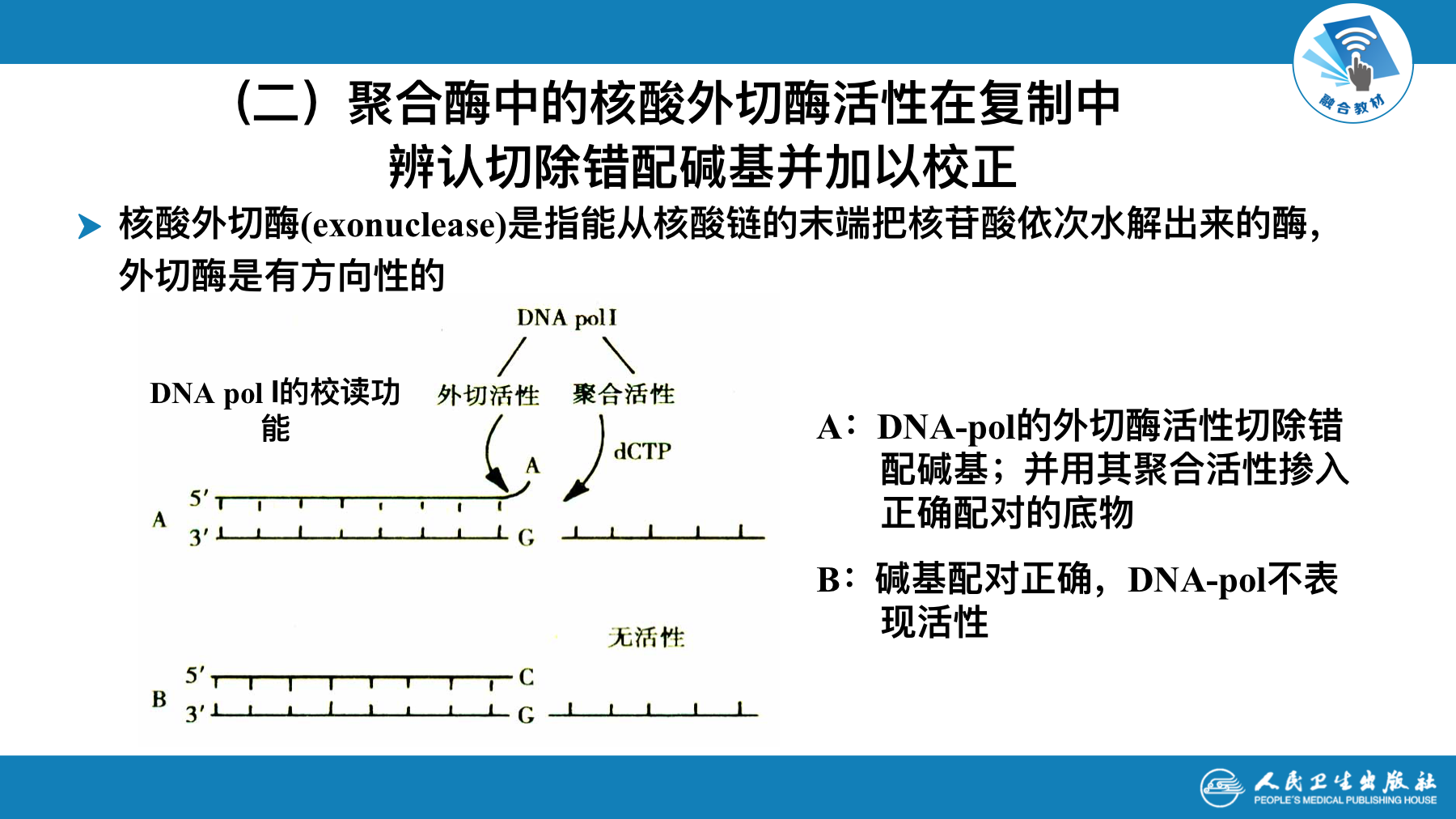
聚合酶的核算外切酶活性在复制中辨别切除错配碱基并加以校正
复制DNA分子拓扑学变化
DNA分子的碱基埋在双螺旋内部,只有把DNA解成单链,它才能起模板作用。
多种酶参与DNA解链和稳定单链状态
原核生物复制中参与DNA解链的相关蛋白质
| 蛋白质(基因) | 通用名 | 功 能 |
|---|---|---|
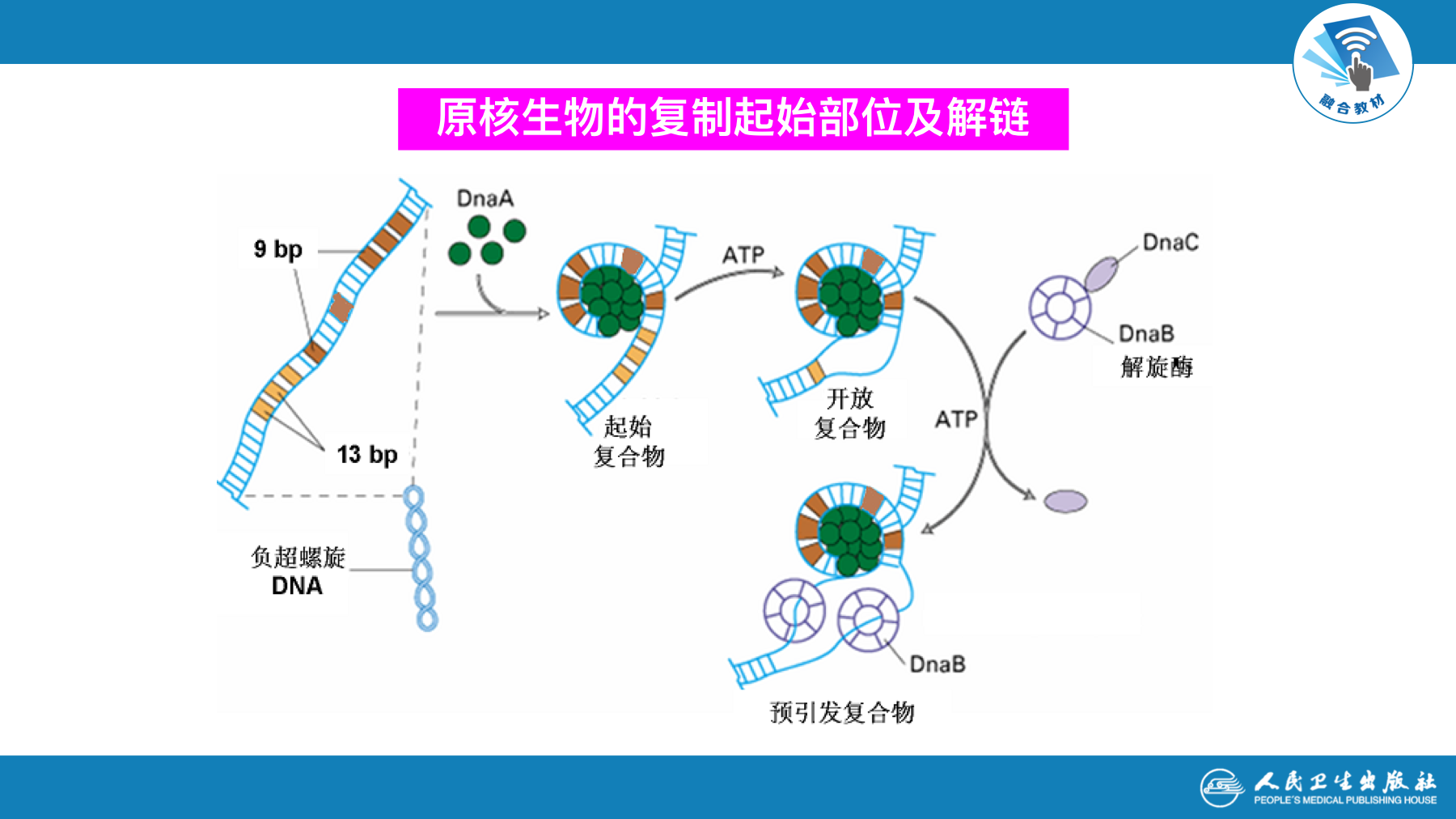
| DnaA(dnaA) | 辨认复制起始点 | |
| DnaB(dnaB) | 解旋酶 | 解开DNA双链 |
| DnaC(dnaC) | 运送和协同DnaB | |
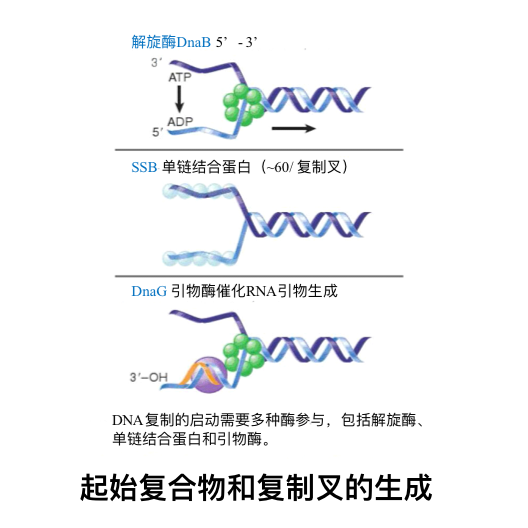
| DnaG(dnaG) | 引发酶 | 催化RNA引物生成 |
| SSB | 单链结合蛋白/DNA结合蛋白 | 稳定已解开的单链DNA |
| 拓扑异构酶 | 拓扑异构酶Ⅱ又称促旋酶 | 解开超螺旋 |
相关酶的参与
- 解螺旋酶(helicase) ——利用ATP供能,作用于氢键,使DNA双链解开成为两条单链
- 引物酶(primase) ——复制起始时催化生成RNA引物的酶
单链DNA结合蛋白(single stranded DNA binding protein, SSB) ——在复制中维持模板处于单链状态并保护单链的完整性
正超螺旋的形成
正常DNA螺旋为负超螺旋,正超螺旋形成后,会相应的形成负超螺旋抵抗
- 环状正超螺旋-(单链被剪切)->环状-(完整剪切)->链状
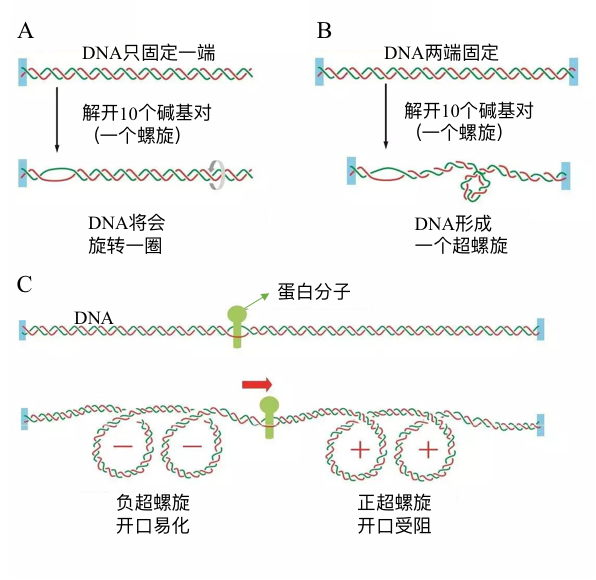
拓扑异构酶
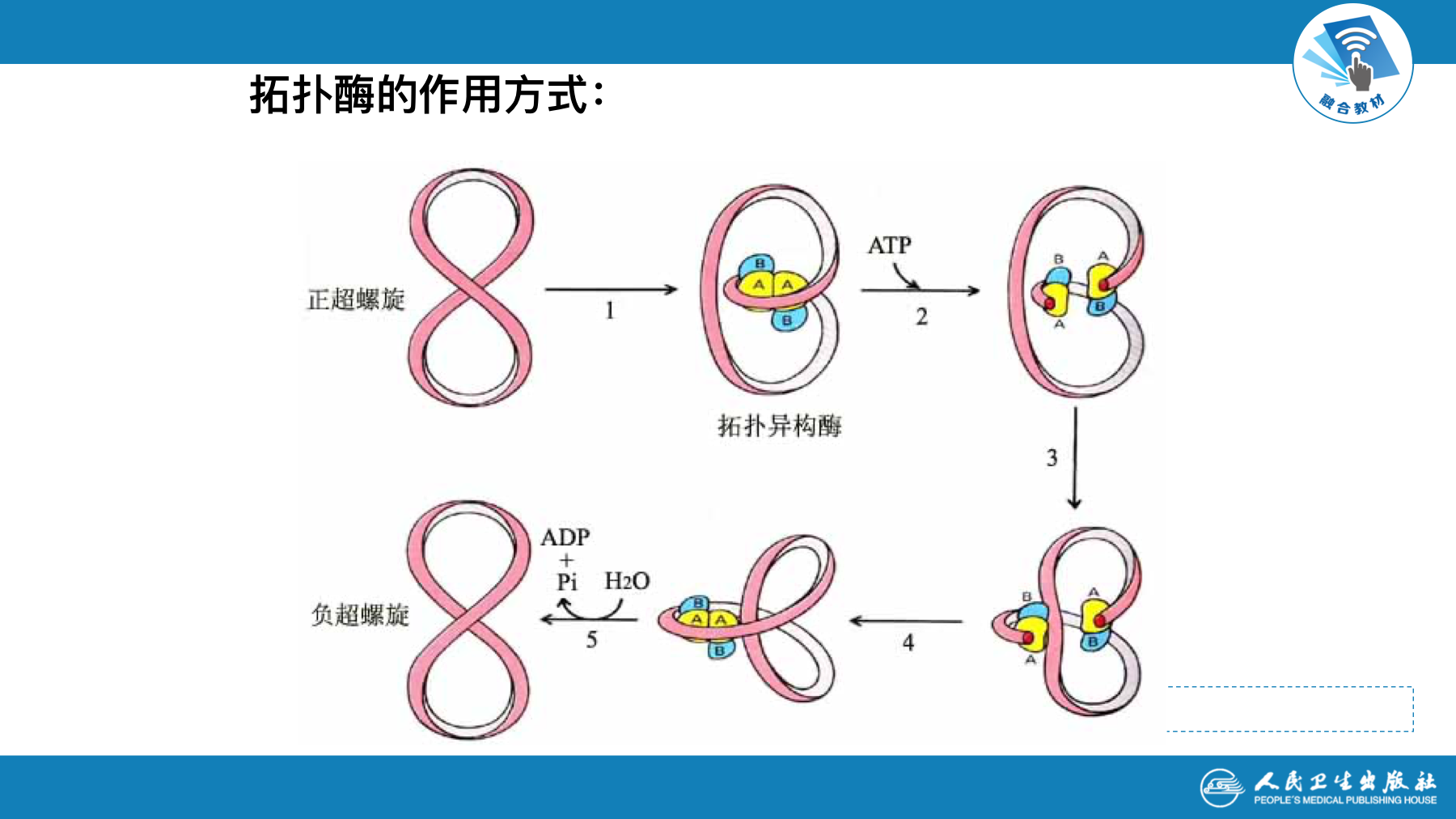
- 拓扑异构酶作用特点:
- 既能水解 、又能连接磷酸二酯键
- 拓扑异构酶分类及作用机制
- 拓扑异构酶Ⅰ:切断DNA双链中一股链,使DNA解链旋转不致打结;适当时候封闭切口,DNA变为松弛状态。反应不需ATP
- 拓扑异构酶Ⅱ:切断DNA分子两股链,断端通过切口旋转使超螺旋松弛利用ATP供能,连接断端, DNA分子进入负超螺旋状态
DNA连接酶连接复制中产生的单链切口
DNA连接酶(DNA ligase)作用方式
- 连接DNA链3’-OH末端和相邻DNA链5’-P末端,使二者生成磷酸二酯键,从而把两段相邻的DNA链连接成一条完整的链
- 通常只作用在单链
- 功能
- DNA连接酶在复制中起最后接合缺口的作用
- 在DNA修复、重组及剪接中也起缝合缺口作用
- 也是基因工程的重要工具酶之一
T4连接酶(T4 ligase)
- 是一种能连接双链的酶,可催化磷酸
原核生物DNA复制过程
- 起始
- 延长
- 终止
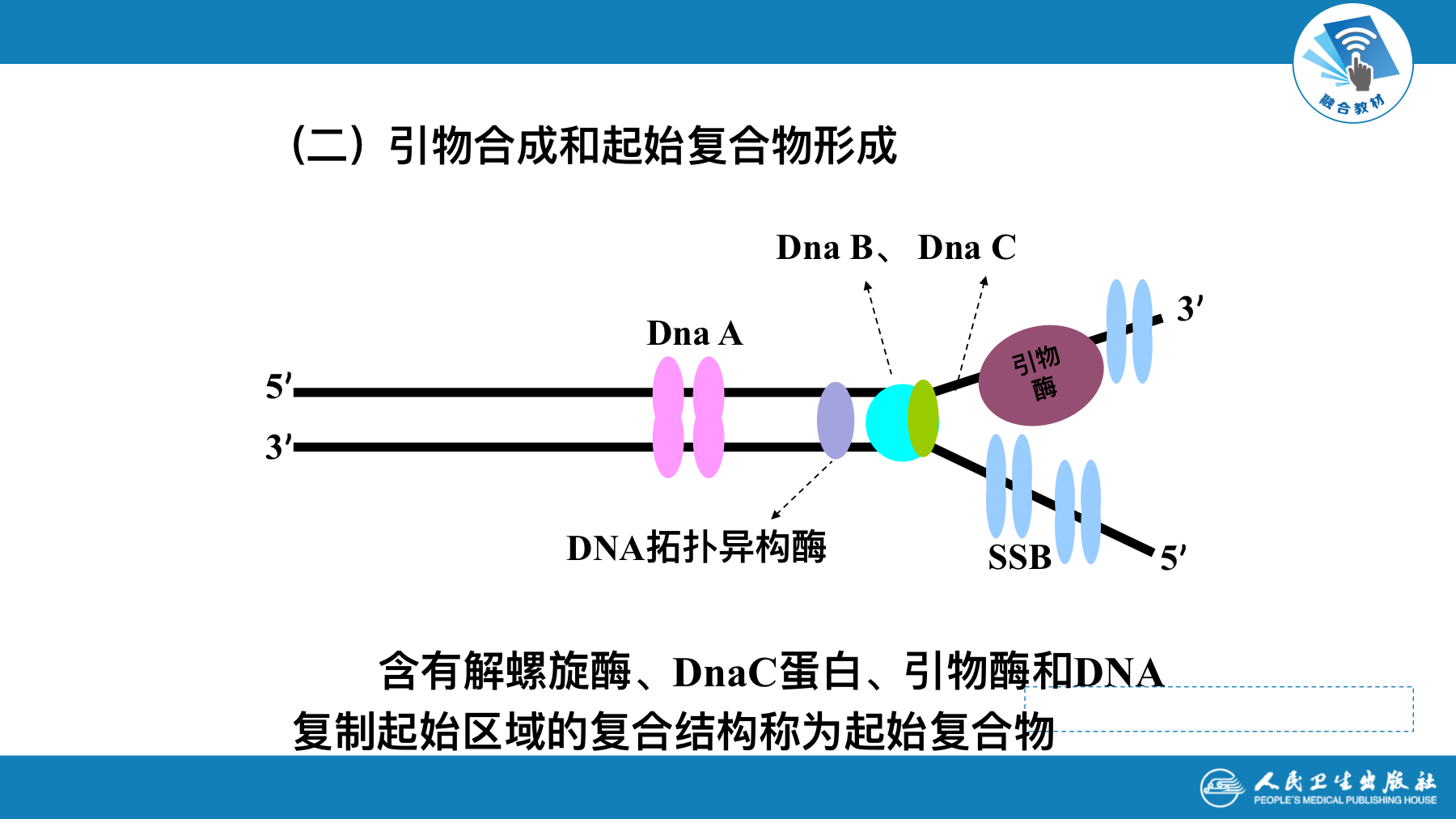
复制的起始
- 起始是复制中较为复杂的环节,在此过程中,各种酶和蛋白因子在复制起始点处装配引发体,形成复制叉并合成RNA引物
- 需要解决两个问题
- DNA解开成单链,提供模版
- 形成引发体,合成引物,提供3’-OH末端
复制的延伸
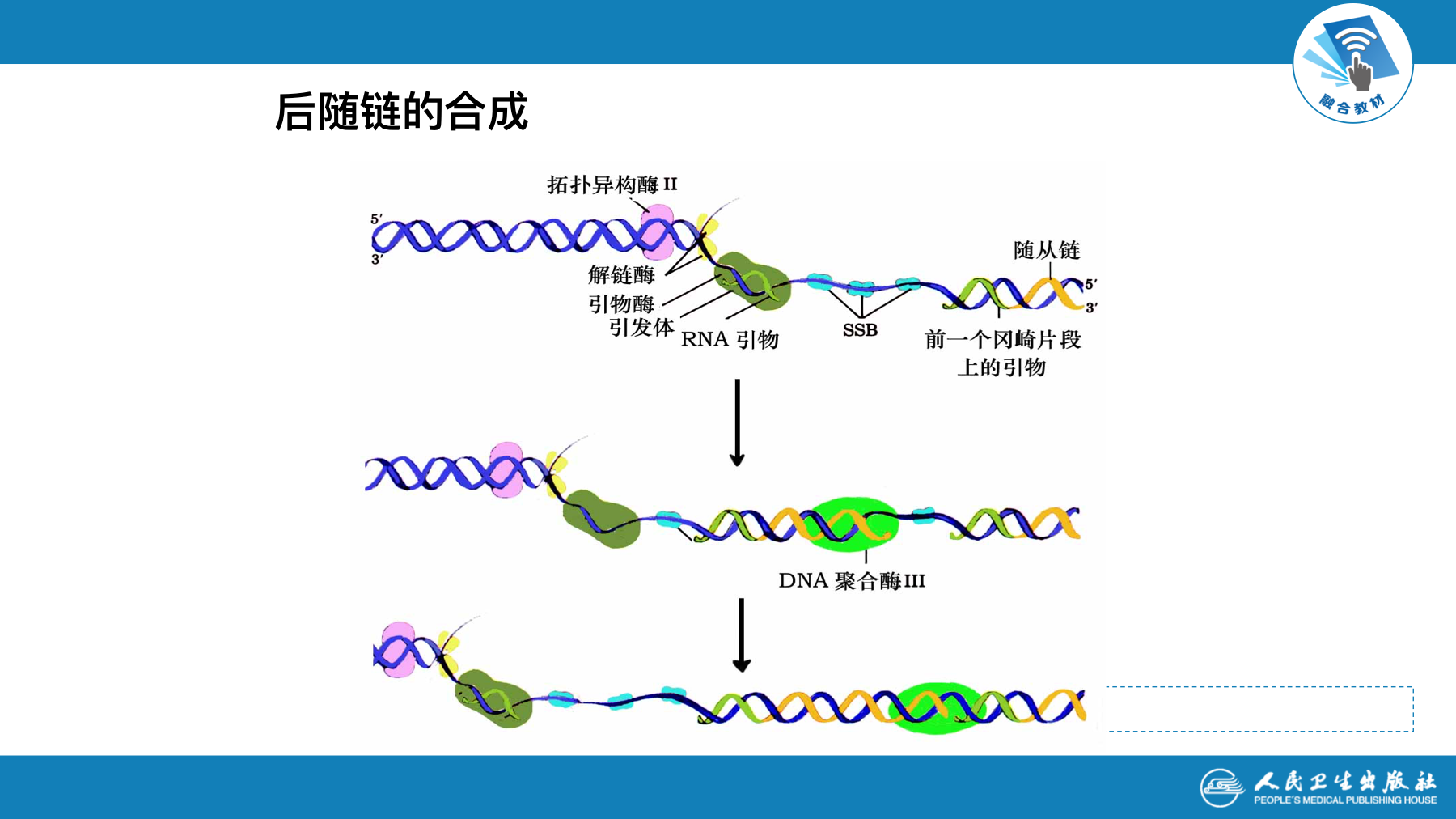
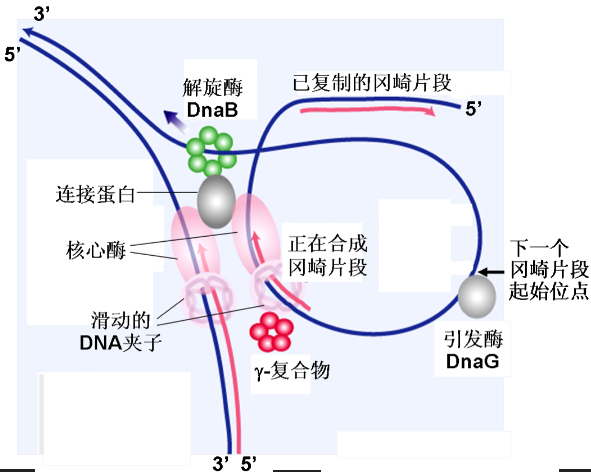
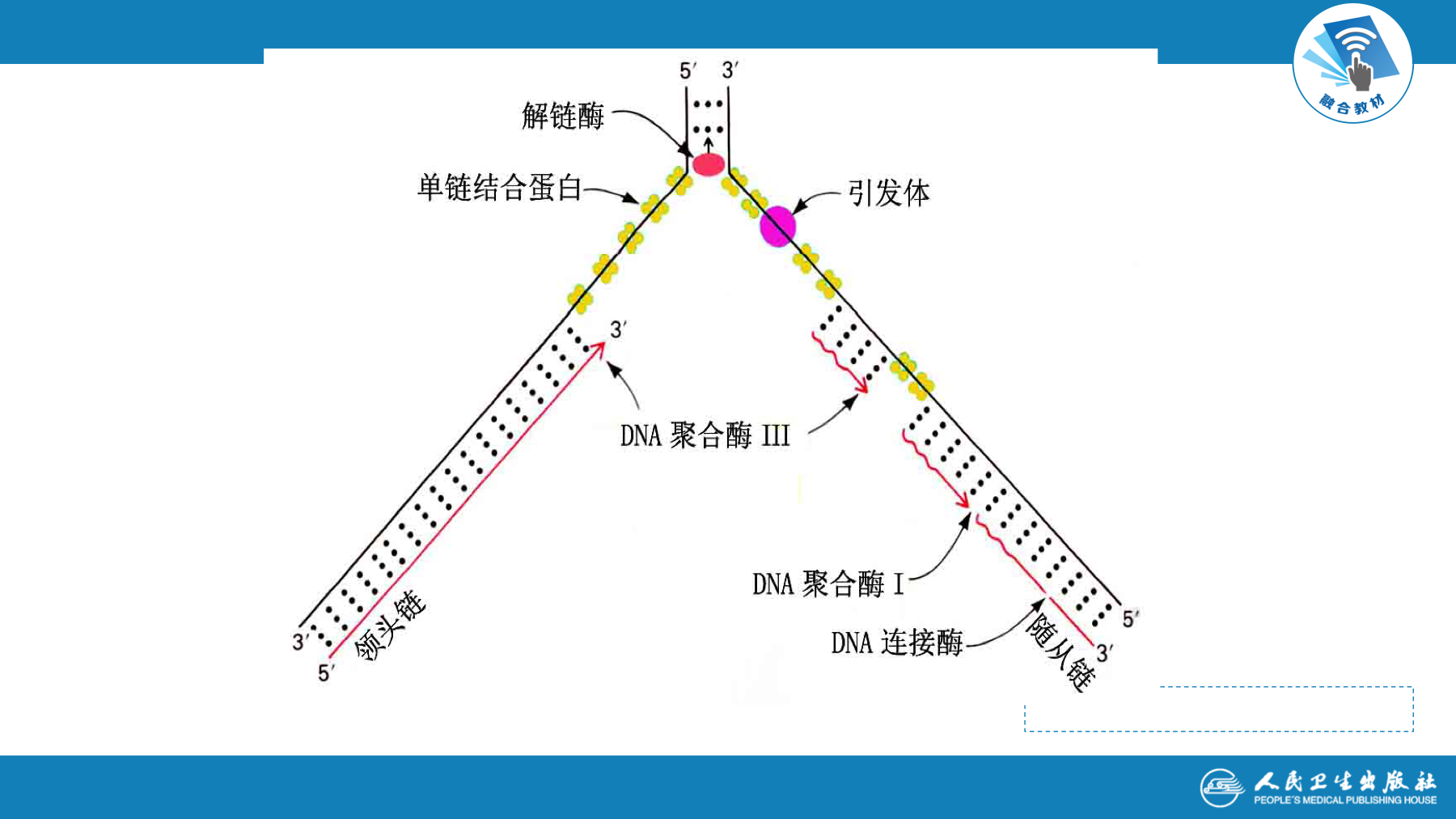
复制的终止
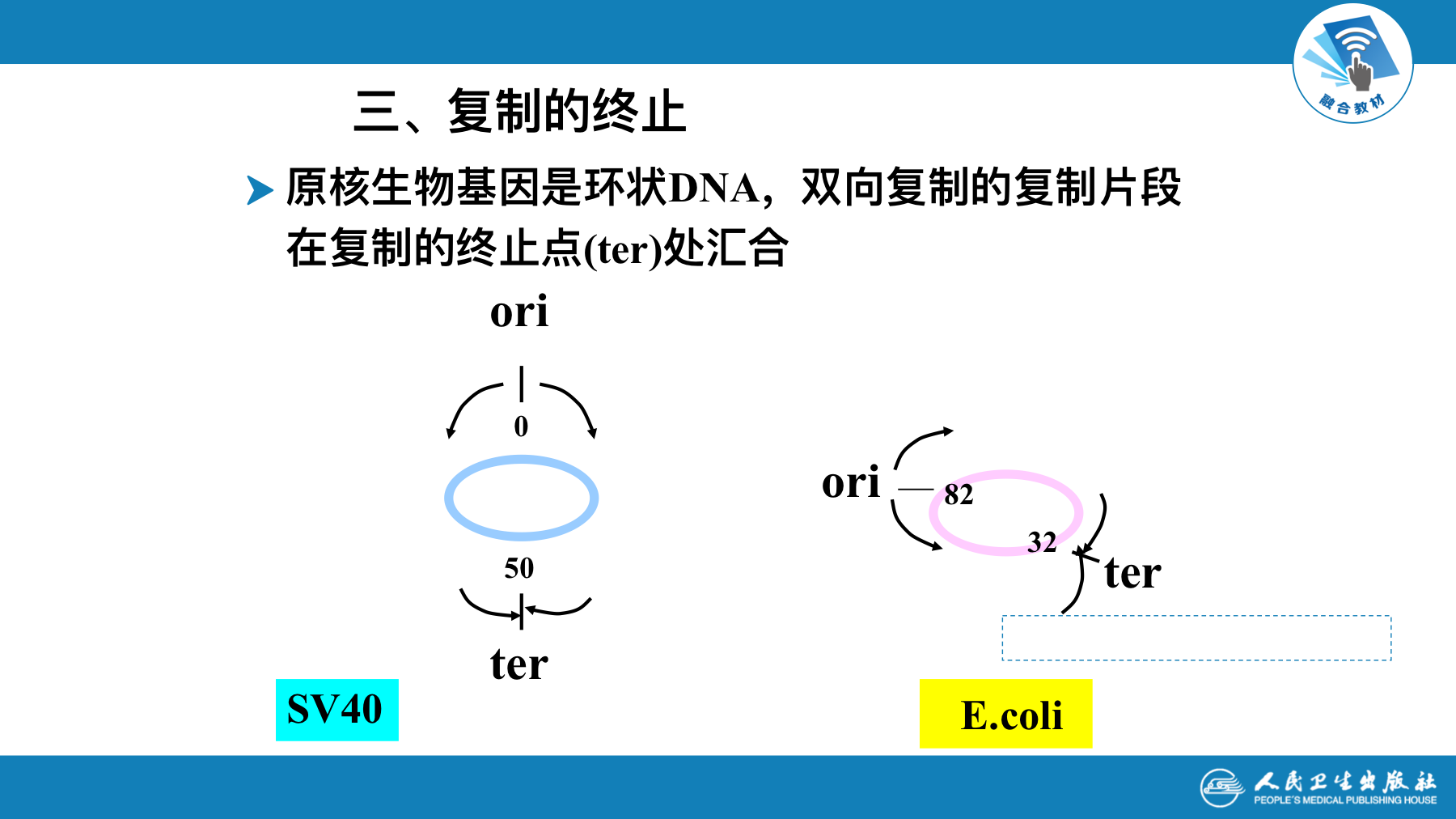
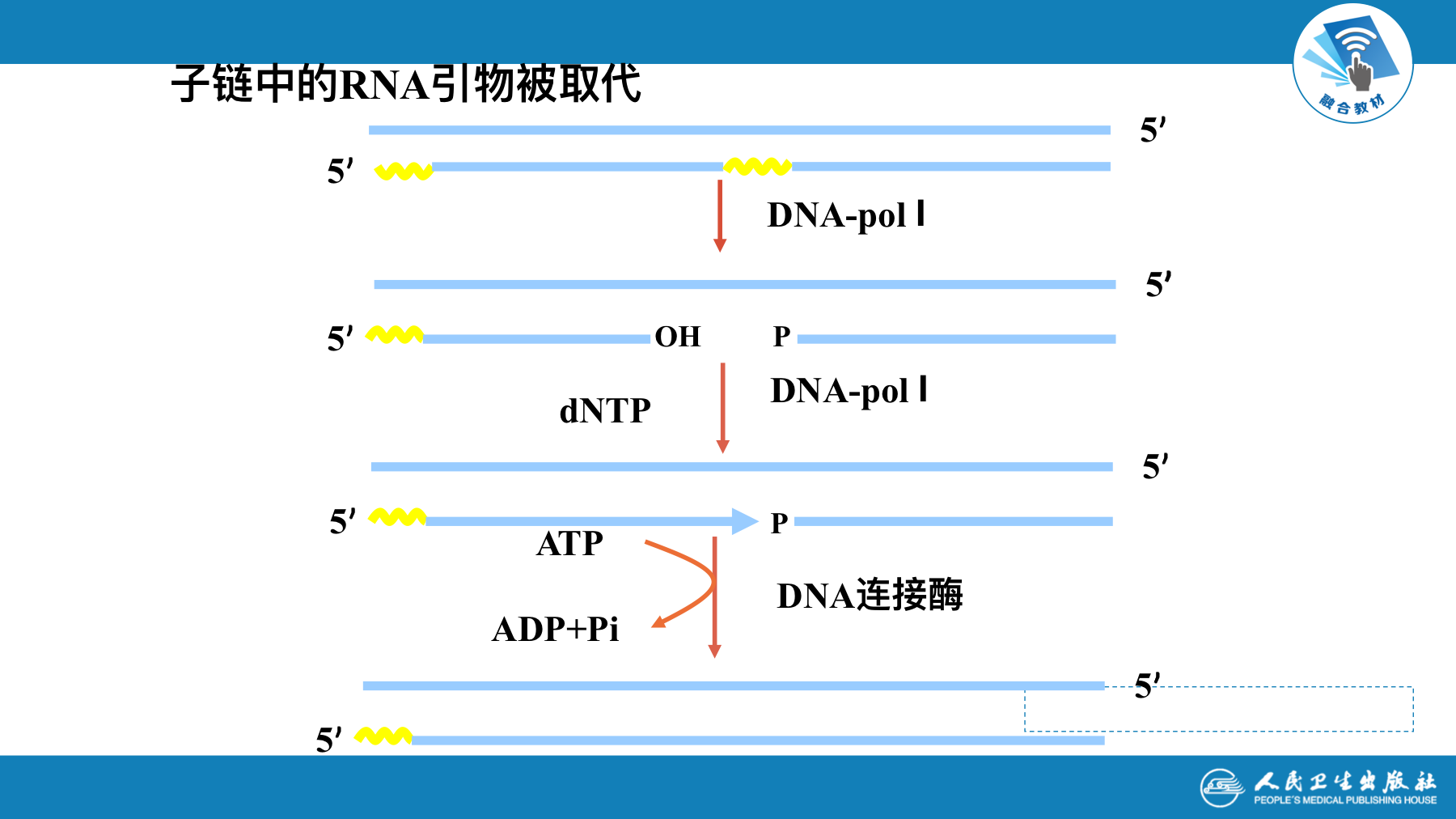
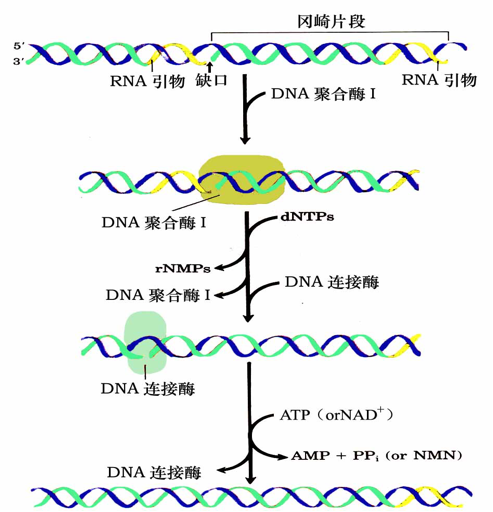
真核生物DNA复制过程
- 真核生物与原核生物DNA复制的差异
- 真核生物复制子多、冈崎片段短、复制叉前进速度慢等
- DNA复制从引发进入延伸阶段发生DNA聚合酶转换
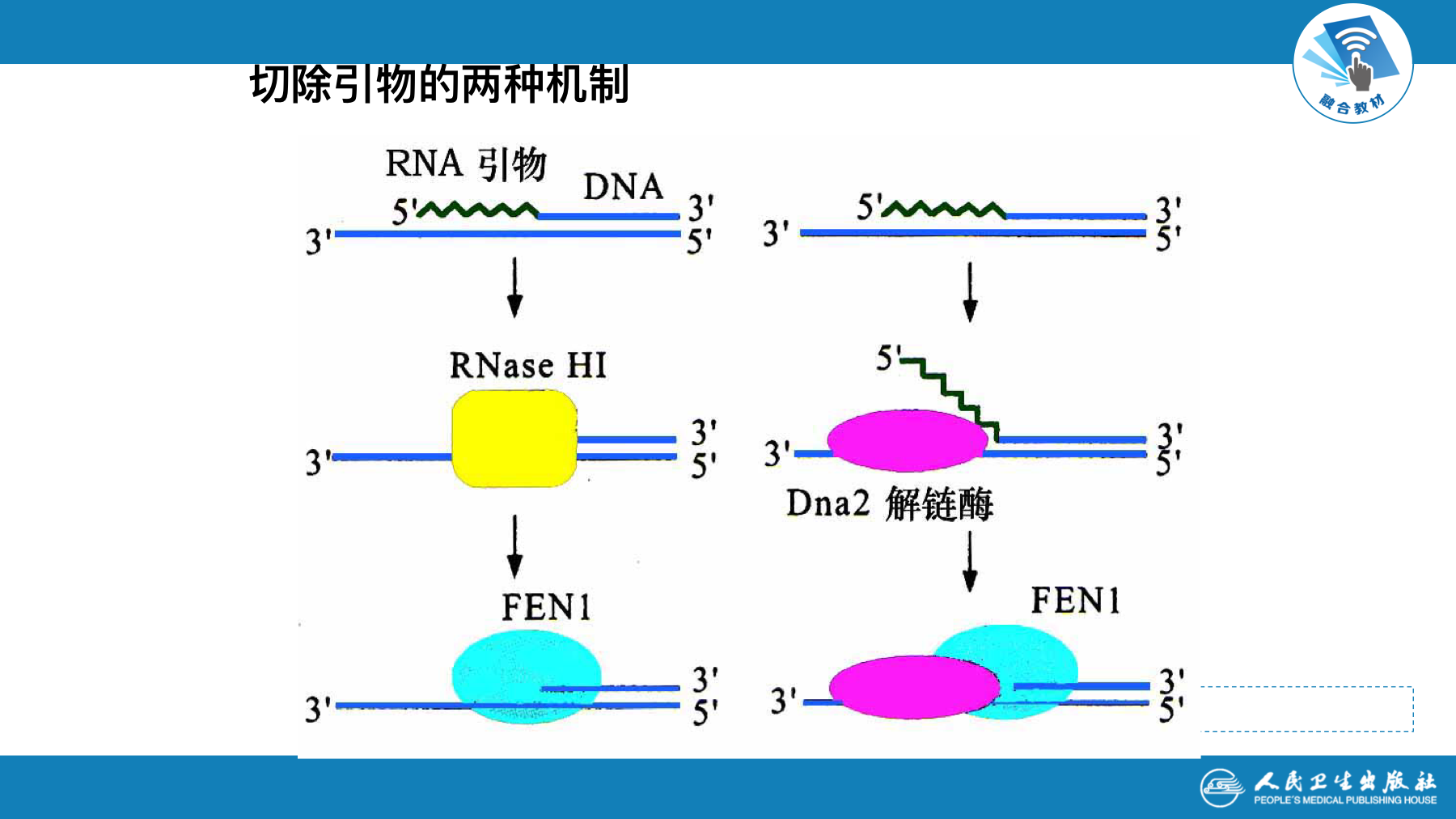
- 切除冈崎片段RNA引物的是核酸酶RNAse H和FEN1等
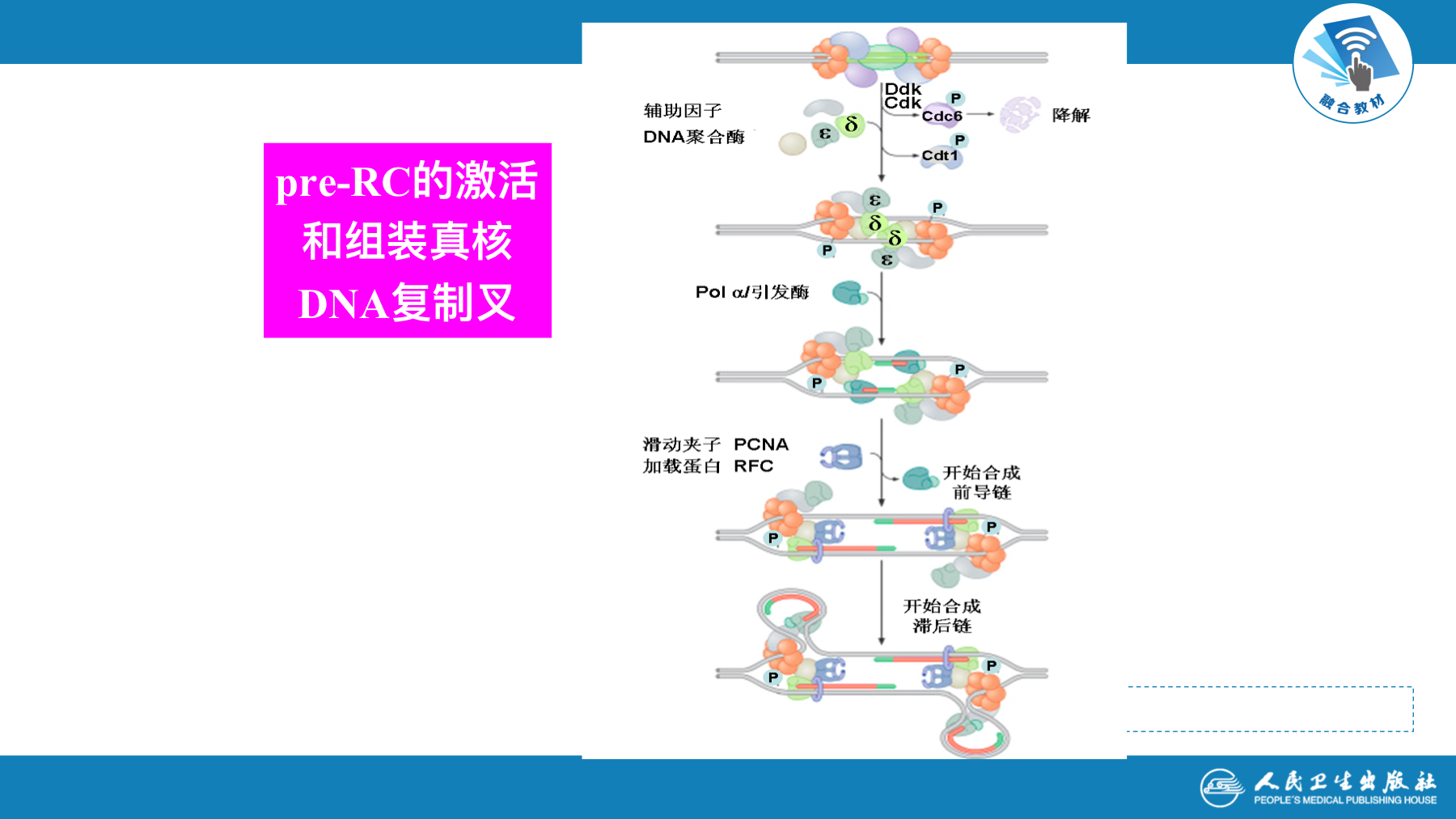
真核生物复制的起始与原核基本相似
- 真核生物每个染色体有多个起始点,是多复制子复制。复制有时序性,即复制子以分组方式激活而不是同步启动
- 复制的起始需要DNA-pol α(引物酶活性)、还需解螺旋酶、拓扑酶和复制因子


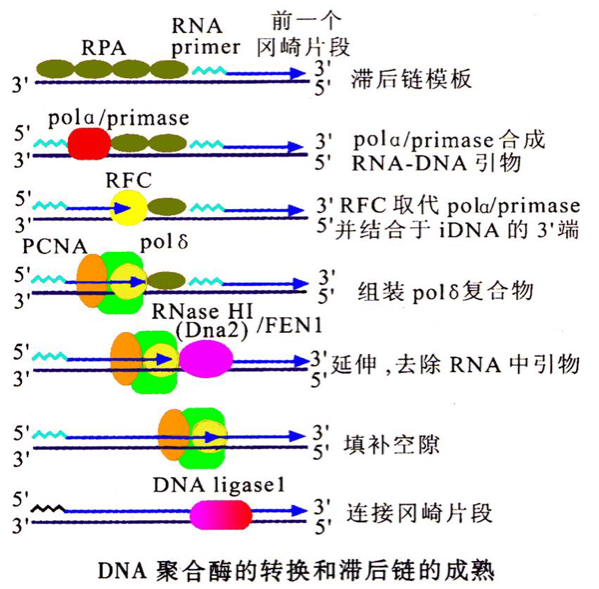
真核生物复制的延长发生DNA聚合酶转换
- 现在认为pol α主要催化合成引物,然后迅速被具有连续合成能力的DNA pol δ和DNA pol ε所替换,这一过程称为聚合酶转换。DNA pol δ负责合成后随链,DNA pol ε负责合成前导链。

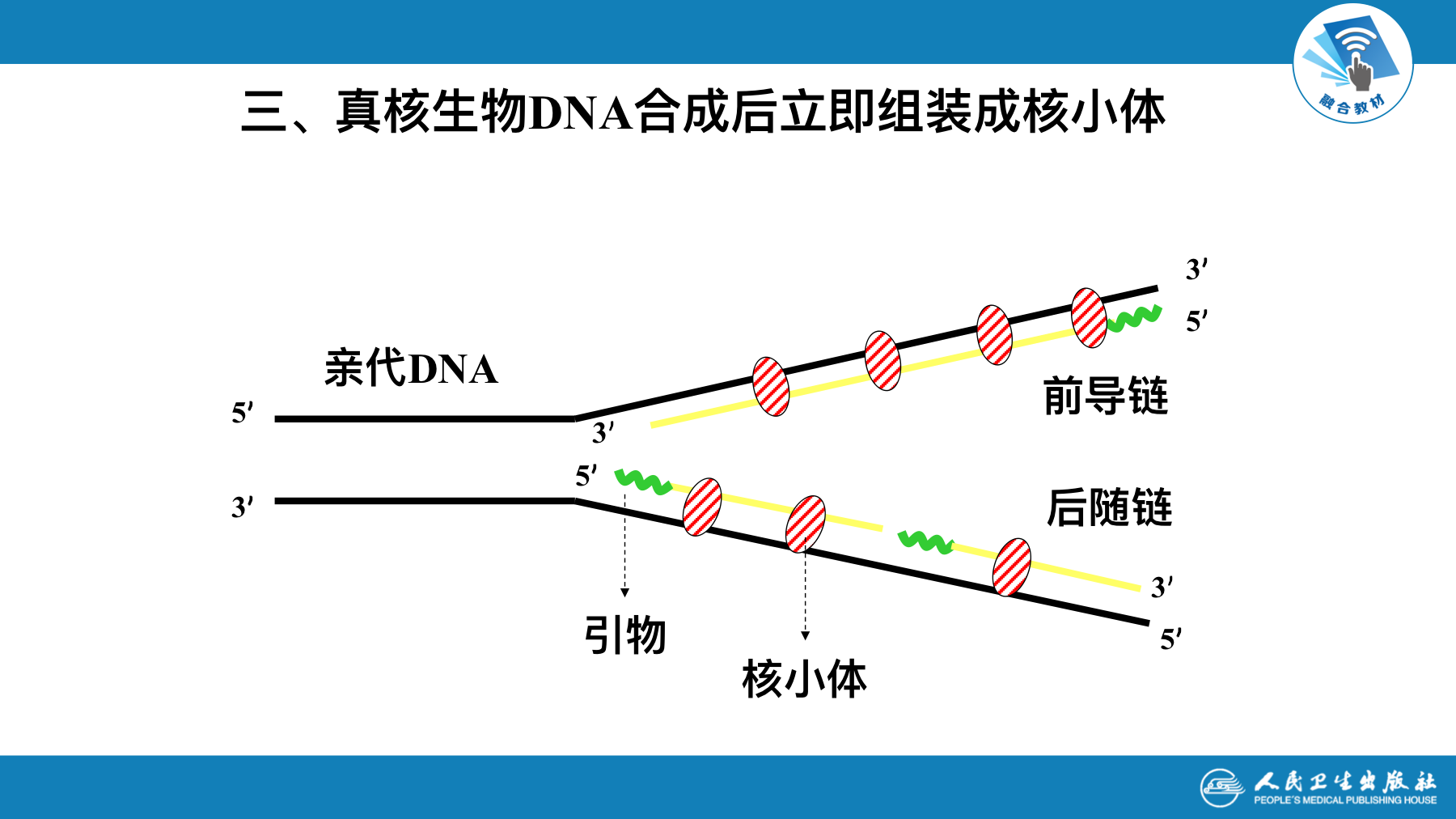
真核生物DNA合成后立即组装成核小体
端粒酶参与解决染色体末端复制问题
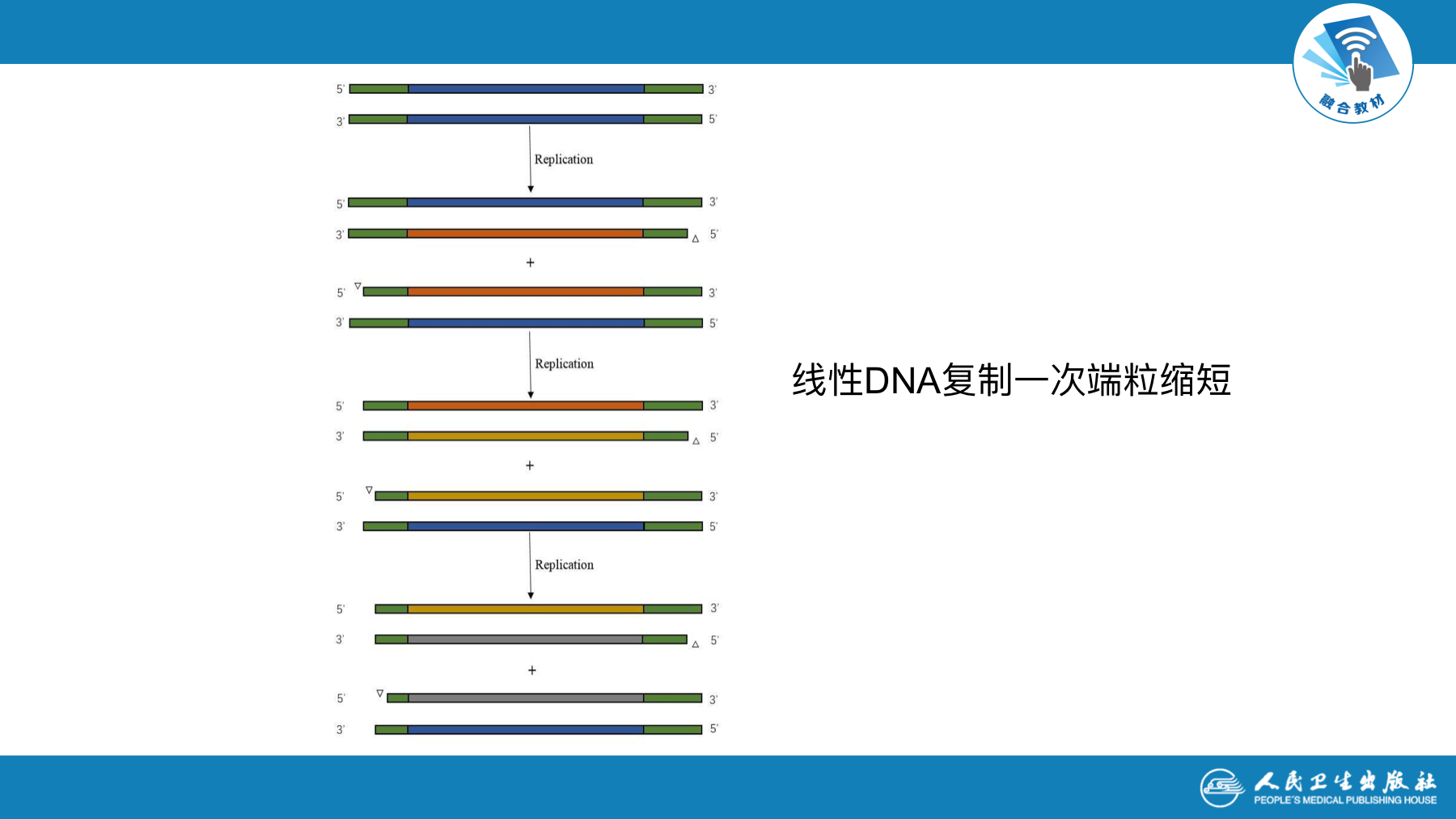
- 染色体DNA呈线状,复制在末端停止
- 复制中冈崎片段的连接,复制子之间的连接
- 染色体两端DNA子链上最后复制的RNA引物,去除后留下空隙
端粒(telomeres)
- 端粒(telomeres)由富含TG的重复序列组成
- 人的端粒重复序列为5’-(TnGn)x-3’ TTAGGG,长为15kb
- 这些重复序列多为双链,但每个染色体的3’端比5’端长,形成ssDNA。这一特殊结构可解决染色体末端复制问题
- 体细胞端粒有限长度(TRFS)大多数明显短于生殖细胞,青年人的TRFs又显著长于年长者,提示TRFs随着细胞分裂衰老
- 端粒末端回折结构
端粒酶(telomerase)
- 组成
- 端粒酶RNA (human telomerase RNA, hTR)
- 端粒酶协同蛋白(human telomerase associated protein 1, hTP1)
- 端粒酶逆转录酶(human telomerase reverse transcriptase, hTRT)
- 端粒酶(telomerase)是一种核糖核蛋白(RNP),由RNA和蛋白质组成
- 端粒酶以自己的RNA组分作为模板,以染色体的3’端ssDNA(后随链模板)为引物,将端粒序列添加于染色体的3’端。这些新合成的DNA为单链

真核生物染色体DNA在每个细胞周期中只能复制一次
- 真核所有染色体DNA复制仅仅出现在细胞周期的S期,而且只能复制一次



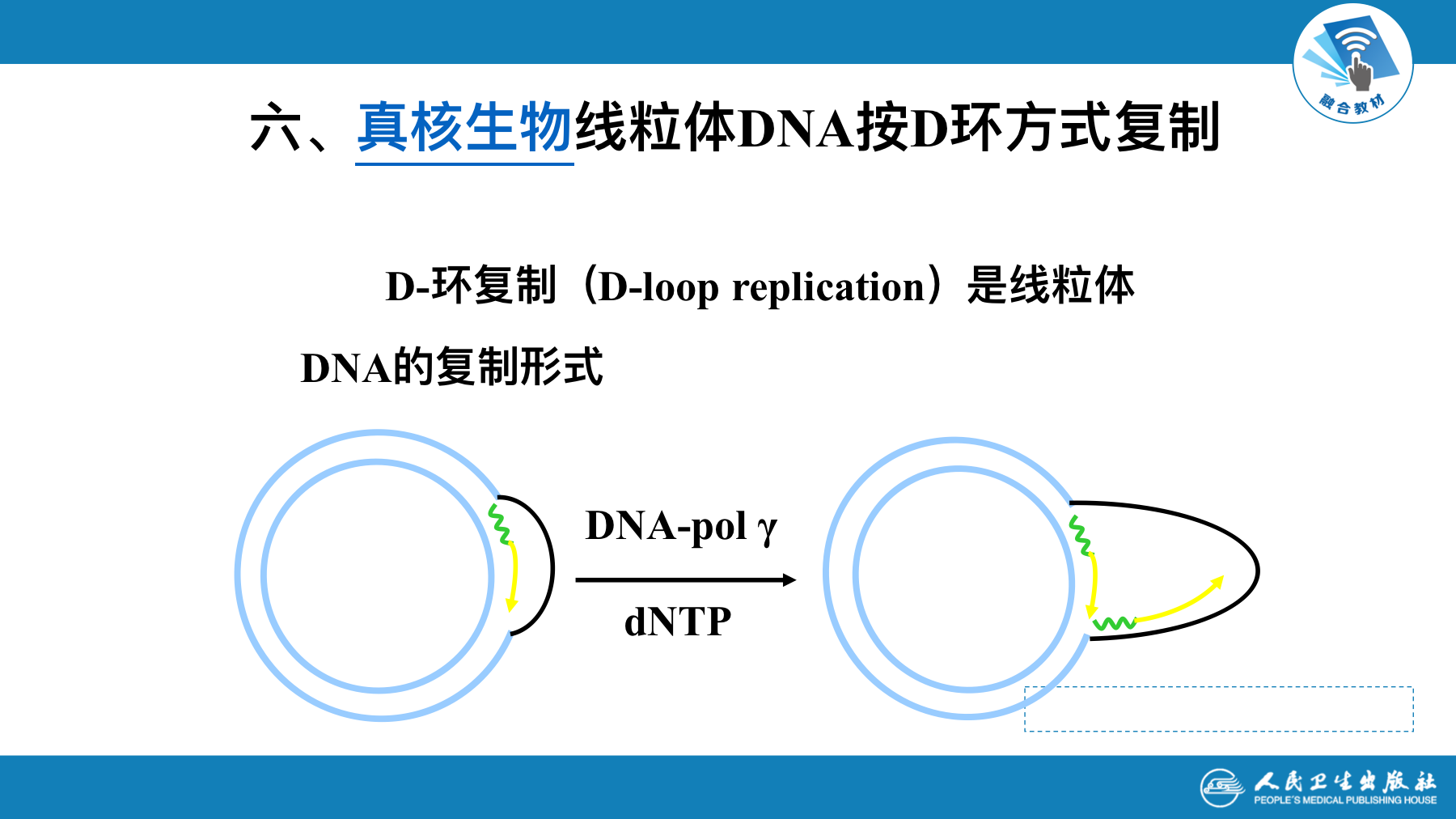
真核生物线粒体DNA按D环方式复制
逆转录
逆转录病毒的基因组以逆转录机制复制
- 逆转录酶中还包括DNA聚合酶、RNA聚合酶、RNA水解酶的活性
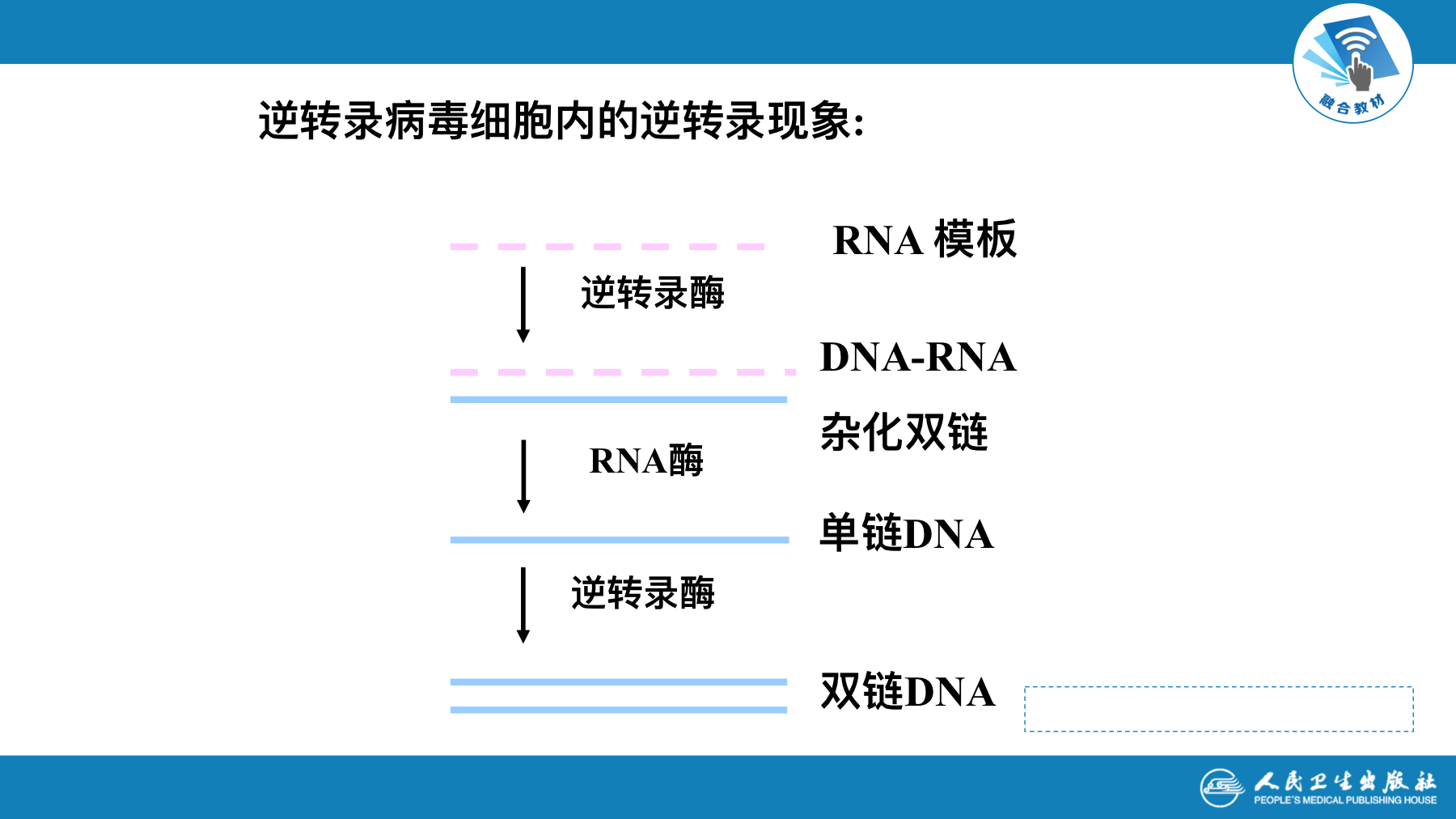
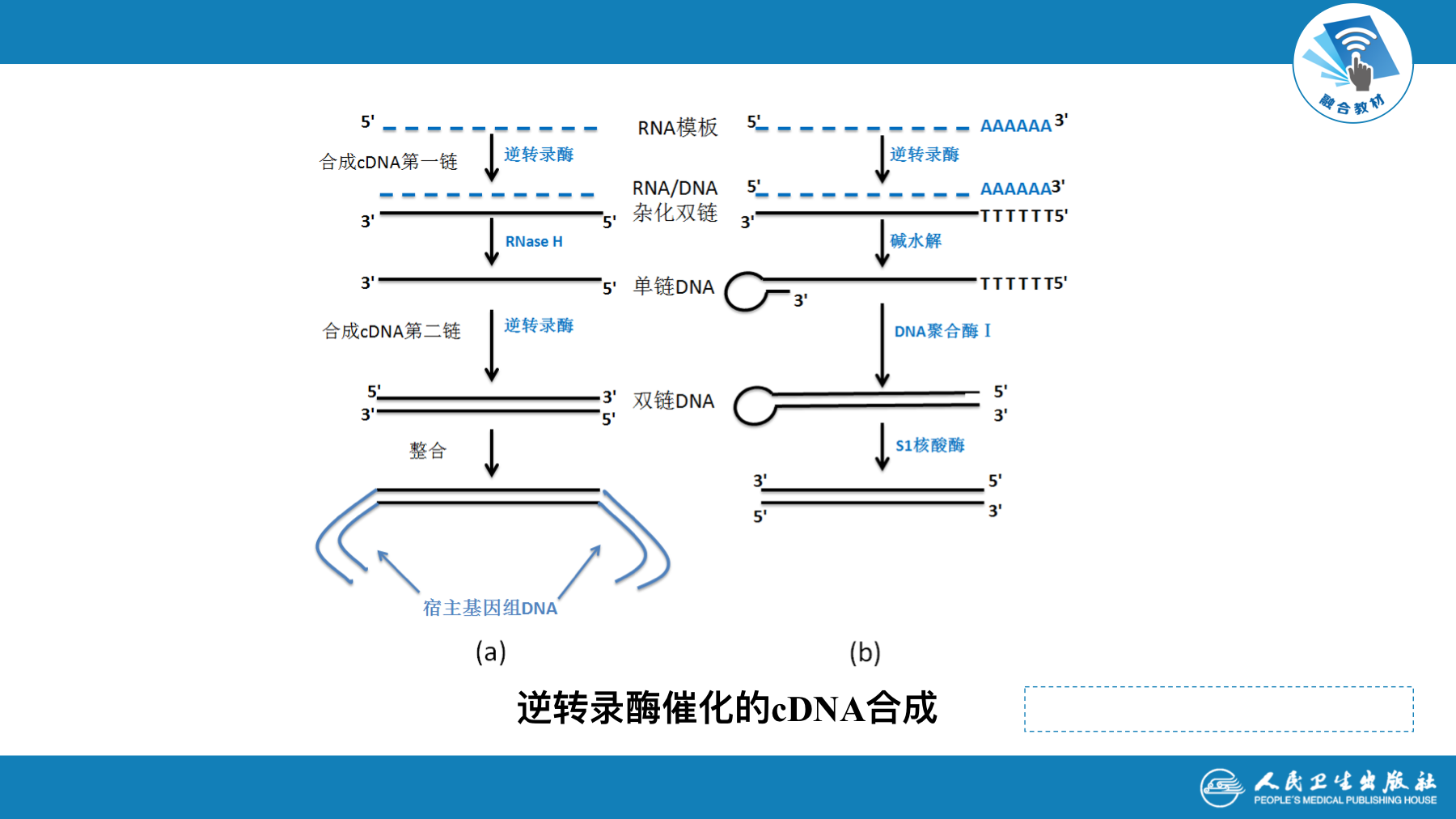
- RNA病毒在细胞内复制成双链DNA的前病毒(provirus)。前病毒保留了RNA病毒全部遗传信息,并可在细胞内独立繁殖。在某些情况下,前病毒基因组通过基因重组(recombination),参加到细胞基因组内,并随宿主基因一起复制和表达。这种重组方式称为整合(integration)。前病毒独立繁殖或整合,都可成为致病的原因
- cDNA-逆转录DNA
能形成磷酸二酯键的酶
| 酶名 | 酶作用 |
|---|---|
| DNA聚合酶 | 以dNTP为底物,遵照碱基互补配对原则,以DNA单链为模版按碱基互补配对原则从5’到3’端通过磷酸二酯键合成子链DNA,起DNA聚合作用 |
| RNA聚合酶 | 是以一条DNA链或RNA为模板,三磷酸核糖核苷为底物、通过磷酸二酯键而聚合的合成RNA的酶 |
| 拓扑异构酶 | 拓扑异构酶Ⅰ:切断DNA双链中一股链,使DNA解链旋转不致打结;适当时候封闭切口,DNA变为松弛状态。反应不需ATP 拓扑异构酶Ⅱ:切断DNA分子两股链,断端通过切口旋转使超螺旋松弛利用ATP供能,连接断端, DNA分子进入负超螺旋状态 |
| 引物酶 | 引物酶是用来引导RNA引物/引子(RNA primer)的合成,从而引导DNA聚合酶介导的DNA链的合成。 |
| 端粒酶 | 端粒酶(telomerase)是一种核糖核蛋白(RNP),由RNA和蛋白质组成。端粒酶以自己的RNA组分作为模板,以染色体的3’端ssDNA(后随链模板)为引物,以磷酸二酯键形式合成单链DNA,并以此将端粒序列添加于染色体的3’端。 |
| DNA连接酶 | 连接DNA链3’-OH末端和相邻DNA链5’-P末端,使二者生成磷酸二酯键,从而把两段相邻的DNA链连接成一条完整的链 |
| 逆转录酶 | 该酶以RNA为模板,以dNTP为底物,tRNA( 主要是色氨酸tRNA) 为引物,tRNA3’-OH 末端上,根据碱基配对的原则,按5’-3’方向以磷酸二酯键方式合成一条与RNA模板互补的DNA单链 |

若有收获,就点个赞吧
0 人点赞
