基因表达调控的基本概念与特点
基因表达产生有功能的蛋白质和RNA
- 基因(gene) 编码蛋白质或RNA等具有特定功能产物的、负载遗传信息的基本单位。
基因表达(gene expression) 基因表达就是基因转录及翻译的过程。也是基因所携带的遗传信息表现为表型的过程,包括基因转录成互补的RNA序列,对于蛋白质编码基因,mRNA继而翻译成多肽链,并装配加工成最终的蛋白质产物。
基因表达具有时间特异性和空间特异性
时间特异性
时间特异性(temporal specificity)
按功能需要,某一特定基因的表达严格按一定的时间顺序发生,称之为基因表达的时间特异性(temporal specificity)。
阶段特异性(stage specificity)
多细胞生物基因表达的时间特异性又称阶段特异性(stage specificity)。
空间特异性
空间特异性(spatial specificity)
在个体生长、发育过程中,一种基因产物在个体的不同组织或器官表达,即在个体的不同空间出现,这就是基因表达的空间特异性(spatial specificity)
细胞特异性(cell specificity)或组织特异性(tissue specificity)
基因表达伴随时间或阶段顺序所表现出的这种空间分布差异,实际上是由细胞在组织器官的分布所决定的,因此基因表达的空间特异性又称细胞特异性(cell specificity)或组织特异性(tissue specificity)。
基因表达的方式存在多样性
基因表达调控(regulation of gene expression)
有些基因产物对生命全过程都是必需的或必不可少的。这类基因在一个生物个体的几乎所有细胞中持续表达,不易受环境条件的影响,或称基本表达。这些基因通常被称为管家基因。
-
基本(或组成性)基因表达(constitutive gene expression)
管家基因的表达称为基本(或组成性)基因表达。
- 基本的基因表达只受启动序列或启动子与RNA聚合酶相互作用的影响,而基本不受其他机制调节。
基本的基因表达水平并非绝对“一成不变”,所谓“不变”是相对的。
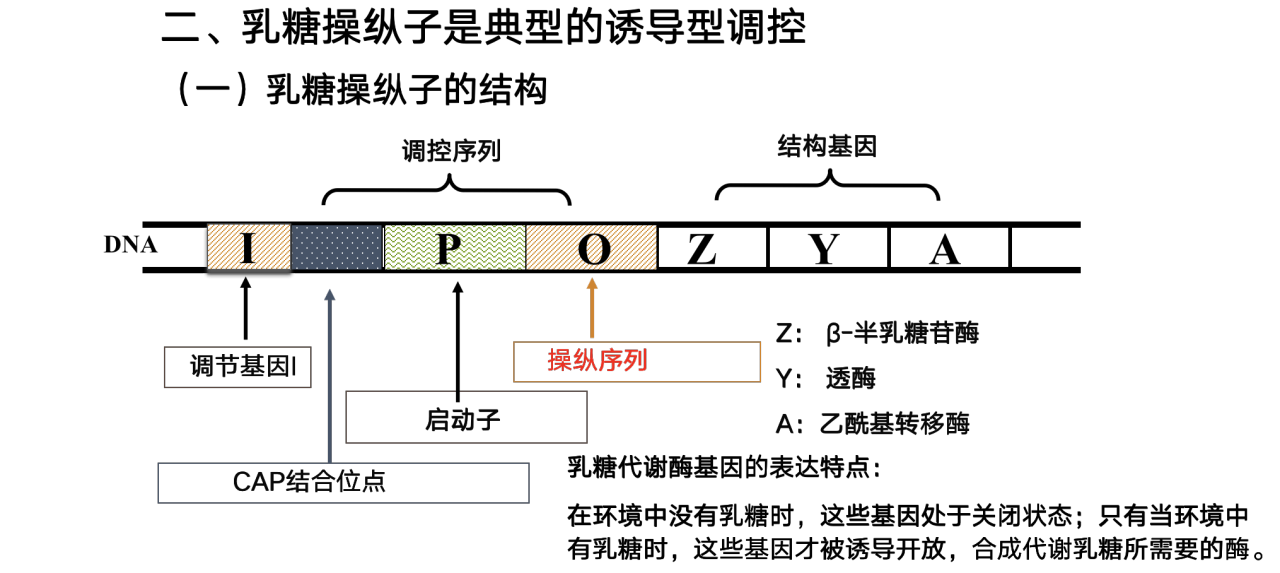
有些基因的表达受到环境变化的诱导和阻遏
-
可诱导基因(inducible gene)
在特定环境信号刺激下,相应的基因被激活,基因表达产物增加,即这种基因表达是可诱导的。例如,在有DNA损伤时,修复酶基因就会在细菌体内被激活,使修复酶被诱导而反应性地增加。
可阻遏基因(repressible gene)
如果基因对环境信号应答时被抑制,这种基因称为可阻遏基因。例如,当培养基中色氨酸供应充分时,细菌体内与色氨酸合成有关的酶编码基因表达就会被抑制。
生物体内不同基因的表达受到协调调节
协同调节(coordinate regulation)
在一定机制控制下,功能上相关的一组基因,无论其为何种表达方式,均需协调一致、共同表达,即为协同表达(coordinate expression)。这种调节称为协同调节。
-
基因表达受调控序列和调节分子的共同调节
顺式作用元件(cis-acting element)
基因的调控序列与被调控的编码序列位于同一条DNA链上,被称为顺式作用元件。
反式作用因子(trans-acting factor)
某些基因的调控序列远离被调控的编码序列,实际上是其他分子的编码基因,只能通过其表达产物来发挥作用,这类调控基因产物称为调节蛋白质。
调节蛋白质不仅能对处于同一条DNA链上的结构基因的表达进行调控,而且还能对不在一条DNA链上的结构基因的表达起到同样的作用。因此,这些蛋白质分子被称为反式作用因子。
基因表达调控呈现多层次和复杂性
染色体水平
染色质结构
- 组蛋白修饰:如乙酰化
- DNA扩增:增加拷贝
- DNA重排:影响转录水平或蛋白质活性
DNA甲基化:CpG岛长常存在于启动子中,C的甲基化抑制转录的启动
转录水平
翻译水平
翻译后加工
原核基因表达调控
原核生物基因组的结构特点
原核生物基因组是具有超螺旋结构的闭合环状DNA分子
原核生物大多数基因表达调控是通过操纵子机制实现的
- 原核生物几个功能相关的基因往往形成表达的协同单位。一群功能相关的结构基因串联在一起,连同其上游的调控区构成一个转录单位,称为操纵子(operon) 。
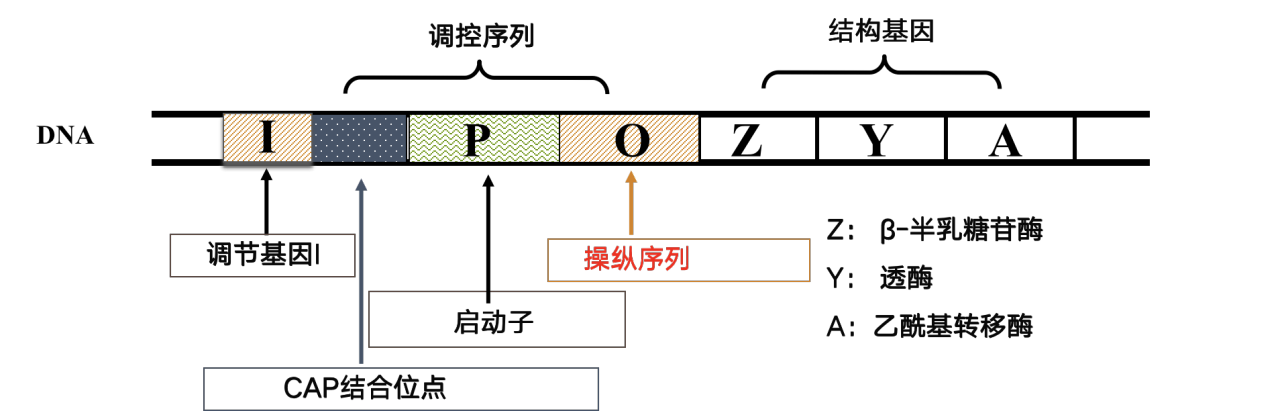
操纵子(operon):由结构基因、调控序列和调节基因组成
- 结构基因:包括数个功能上有关联的基因,它们串联排列,共同构成编码区。这些结构基因共用一个启动子和一个转录终止信号序列,因此转录合成时仅产生一条mRNA长链,为几种不同的蛋白质编码。这样的mRNA分子携带了几个多肽链的编码信息,被称为多顺反子(polycistron)mRNA。
调控序列:包括启动子(promoter)和操纵元件(operator)
启动子:RNA聚合酶和各种调控蛋白作用的部位,是决定基因表达效率的关键元件。
通常在转录起始点上游-10及-35区域存在一些相似序列,称为共有序列。E.coli 及一些细菌启动序列的共有序列在-10区域是TATAAT,又称Pribnow盒,在-35区域为TTGACA。这些共有序列决定启动子的转录活性大小。
操纵元件:是一段能被特异的阻遏蛋白识别和结合的DNA序列。
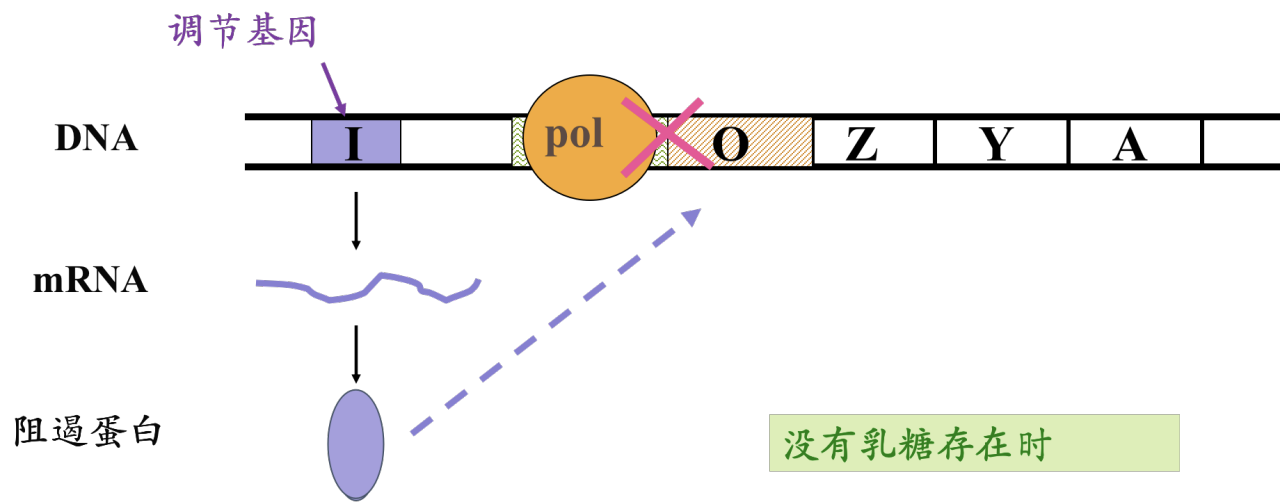
- 调节基因(regulatory gene):编码能够与操纵序列结合的阻遏蛋白
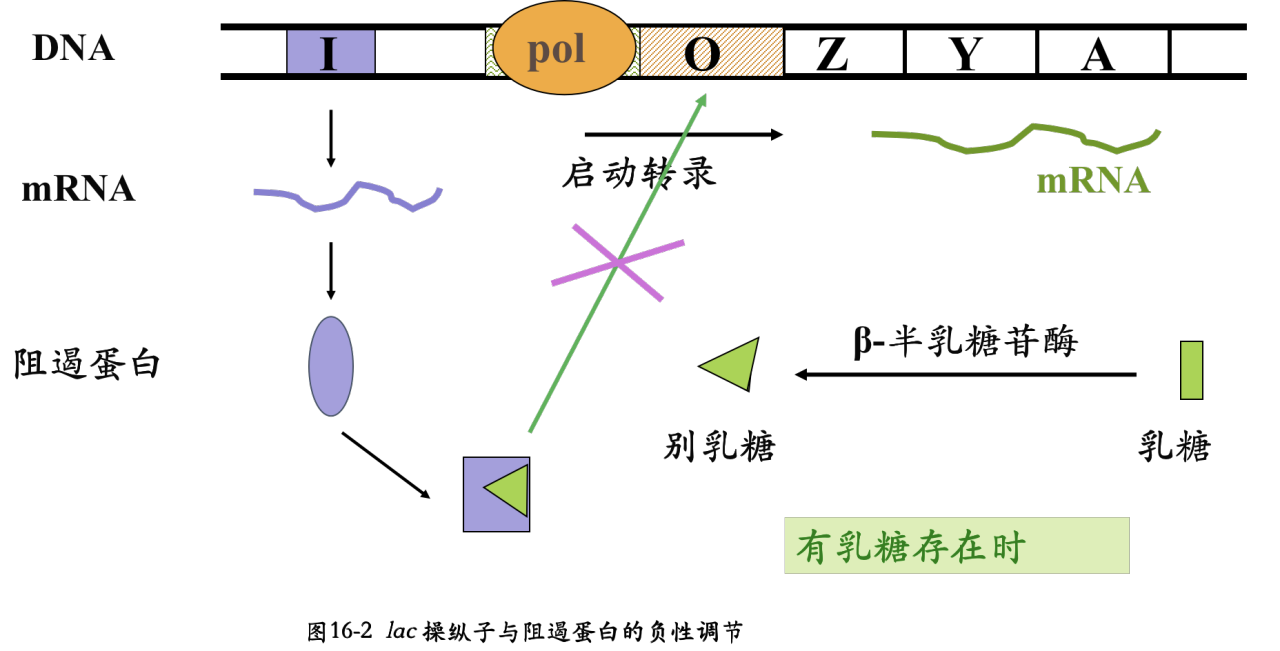
- 阻遏蛋白的作用: 识别、结合特异的操纵序列,抑制基因转录,所以阻遏蛋白介导负调节(negative regulation)。阻遏蛋白介导的负性调节机制在原核生物中普遍存在。
其它调控蛋白:特异因子,激活蛋白
正调节物: CAP。 CAP发挥作用时需要cAMP的参与。 葡萄糖缺乏时,cAMP浓度升高。
- 机理:cAMP与CAP结合形成复合物,促使CAP结合CAP位点,激活RNA聚合酶
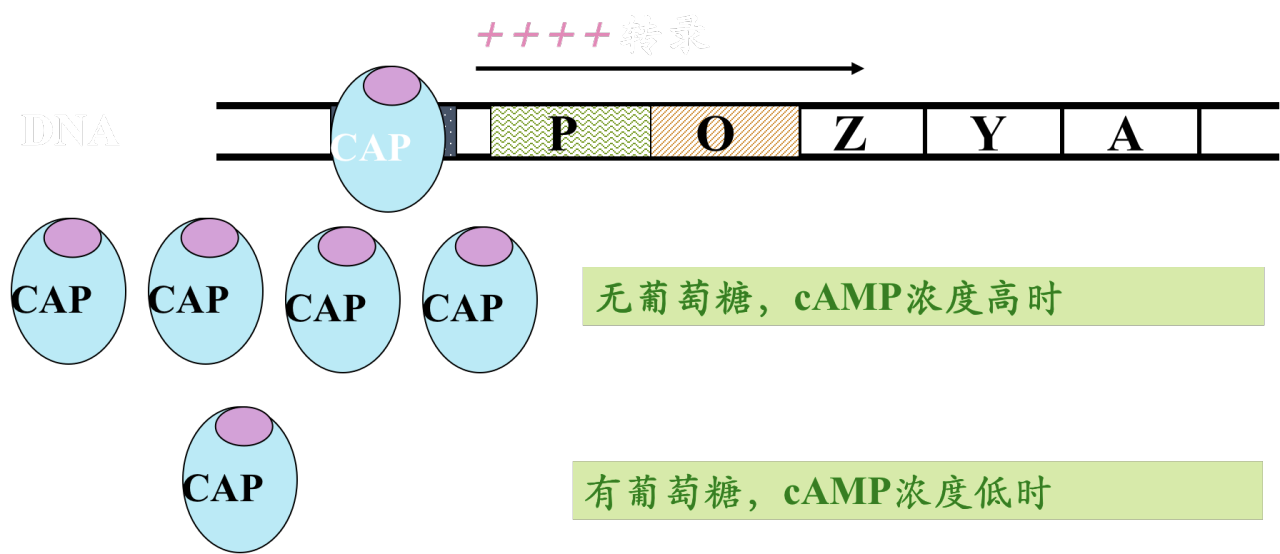
协同调节
- 当阻遏蛋白封闭转录时,CAP对该系统不能发挥作用。
- 如无CAP加强转录活性,即使没有阻遏蛋白与操纵序列结合,操纵子仍几无转录活性。
- 单纯乳糖存在时,细菌利用乳糖作碳源;若有葡萄糖或葡萄糖/乳糖共同存在时,细菌首先利用葡萄糖。葡萄糖对lac操纵子的阻遏作用称分解代谢阻遏(catabolic repression)。
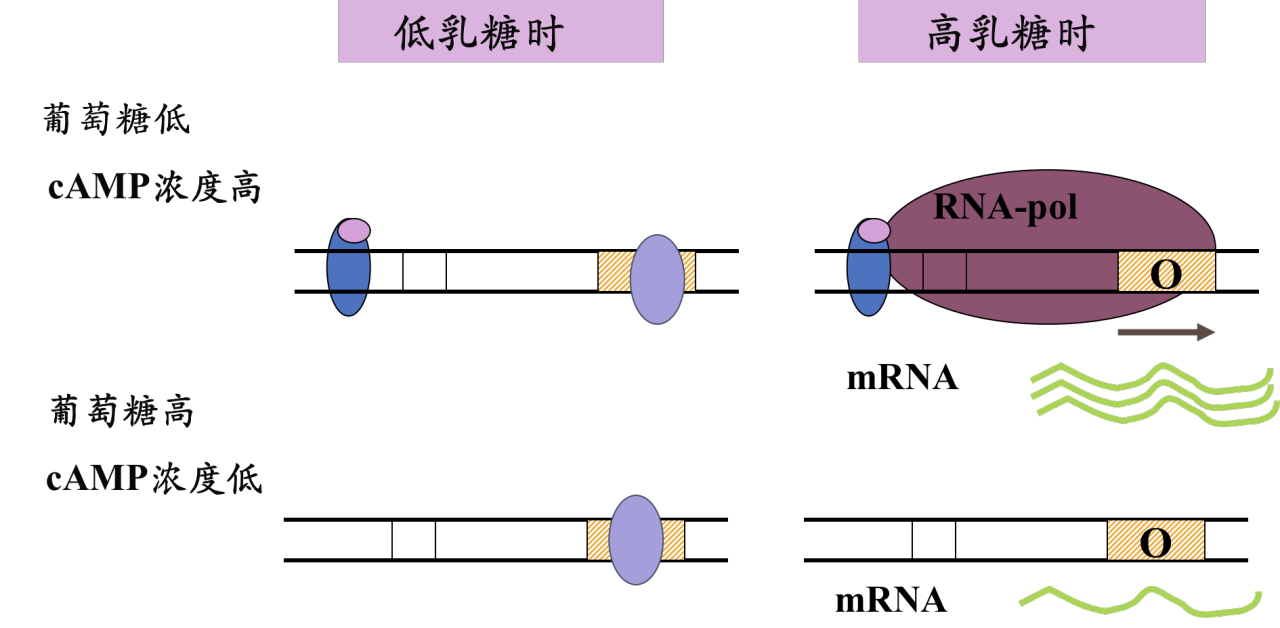
乳糖操纵子既要受到负调控,又要受到正调控的原因
- 使细胞能够优先利用葡萄糖,而优先利用葡萄糖对细胞来说是有益的,因为参与葡萄糖分解的基因均是管家基因,这样葡萄糖可以迅速地被分解,为细胞提供能量
lac启动子序列和启动子序列的一致序列相差较大,是一个小弱启动子,而CAP-cAMP的激动就弥补了其启动子的“先天不足”
色氨酸操纵子通过阻遏作用和转录衰减作用抑制基因表达
色氨酸操纵子通过阻遏作用抑制基因表达
细胞内无色氨酸时,阻遏蛋白不能与O序列结合,因此色氨酸操纵子处于开放状态,结构基因得以表达。
- 细胞内色氨酸的浓度较高时,色氨酸作为辅阻遏物与阻遏蛋白形成复合物并结合到O序列上,关闭色氨酸操纵子,停止表达用于合成色氨酸的各种酶。
-
色氨酸操纵子
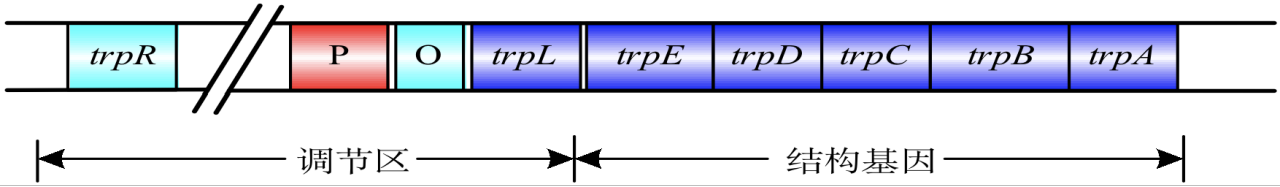
结构基因:trpE、trpD、trpC、trpB 和 trpA
- 上游调控区:调节基因(trpR)、启动子(P)、前导序列(trpL)和操纵序列(O)。
-
开放状态的色氨酸操纵子
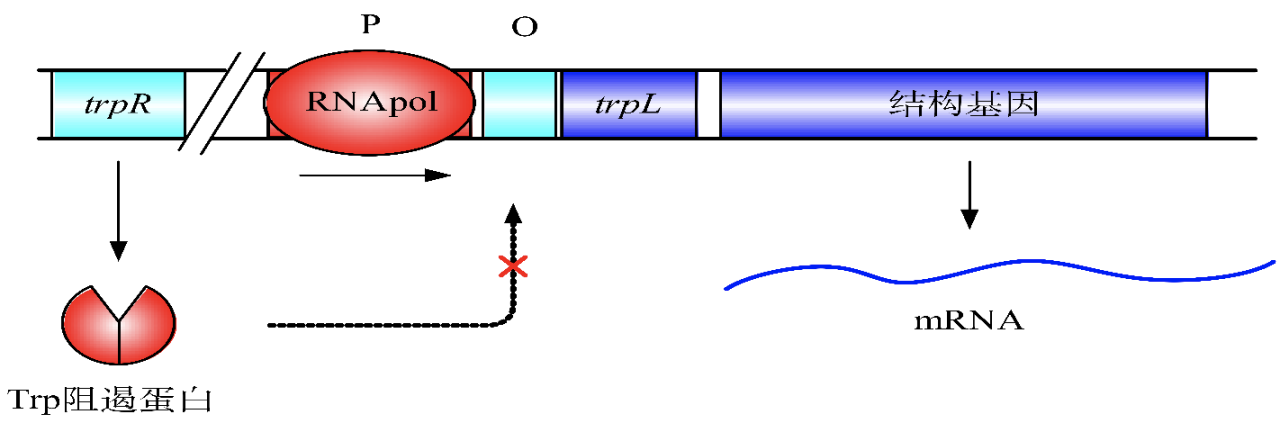
当培养基中色氨酸含量很少时,trpR 阻遏蛋白以同源二聚体的形式存在,不能与操纵序列 O 结合,使得RNA聚合酶能够启动转录。
关闭状态的色氨酸操纵子
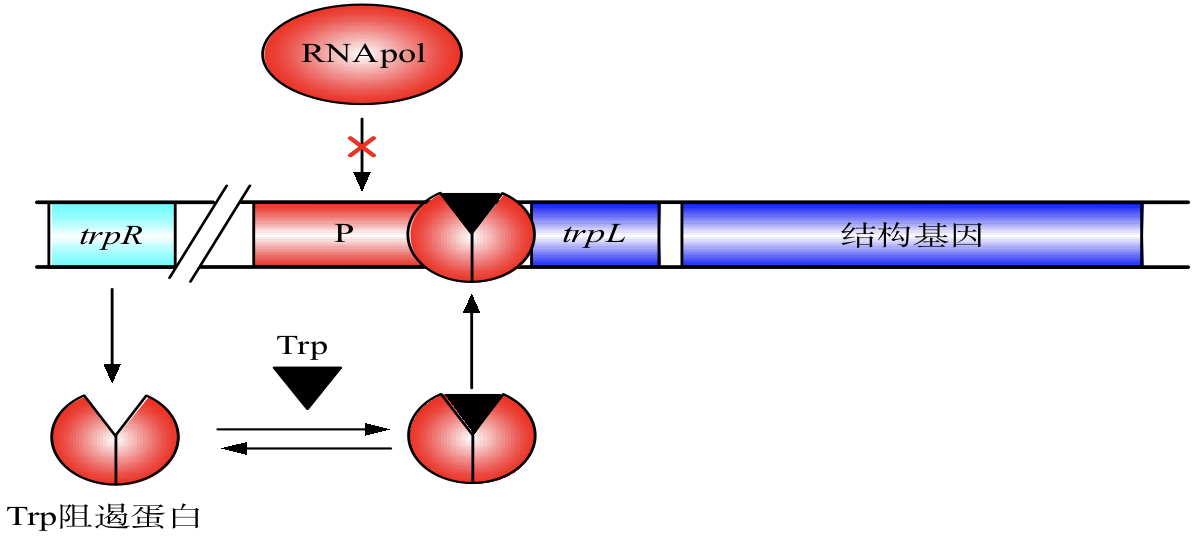
当色氨酸含量丰富时,色氨酸与色氨酸阻遏蛋白结合,使其能够与操纵序列结合,抑制转录。
色氨酸操纵子通过转录衰减作用抑制基因表达
转录衰减(transcription attenuation):使已经开始转录的mRNA合成终止的基因表达调节方式,称为转录衰减。这种作用是利用原核生物中转录与翻译过程偶联进行,翻译时先合成的一段前导序列L来实现的。
前导序列L的结构特点
色氨酸的浓度较低时,前导肽的翻译因色氨酸量的不足而停滞在第10/11的色氨酸密码子部位,核糖体结合在序列1上, 因此前导mRNA倾向于形成2/3发夹结构,转录继续进行;
色氨酸的浓度较高时,前导肽的翻译顺利完成,核糖体可以前进到序列2,因此发夹结构在序列3和序列4形成,连同其下游的多聚U使得转录中途终止,表现出转录的衰减。
转录衰减的生理意义
原核生物这种在色氨酸浓度高时,通过阻遏作用(粗调)和转录衰减机制(精调)共同关闭基因表达的方式,保证了营养物质和能量的合理利用。
原核基因在翻译水平受到精细调节
蛋白质分子结合于自身mRNA靶位点进行自我调节
- 翻译阻遏利用蛋白质与mRNA的结合实现对翻译起始的调控
- 反义RNA利用结合mRNA翻译起始部位的互补序列调节翻译起始
- 反义控制(antisense control):有些细菌或病毒,能够转录产生反义RNA。反义RNA含有与特定mRNA翻译起始部位互补的序列,通过与mRNA杂交阻断30S小亚基对起始密码子的识别及与SD序列的结合,抑制翻译起始。这种调节称为反义控制。
- mRNA密码子的编码频率影响翻译速度
真核生物的基因表达
真核细胞的基因表达特点
- 真核基因组比原核基因组大得多。
- 原核基因组的大部分序列都为编码基因,而哺乳类基因组中大约只有10%的序列编码蛋白质、rRNA、tRNA等,其余90%的序列,包括大量的重复序列功能至今还不清楚,可能参与调控。
- 真核生物编码蛋白质的基因是不连续的,转录后需要剪接去除内含子,这就增加了基因表达调控的层次。
- 原核生物的基因编码序列在操纵子中,多顺反子mRNA使得几个功能相关的基因自然协调控制;而真核生物则是一个结构基因转录生成一条mRNA,即mRNA是单顺反子(monocistron),许多功能相关的蛋白、即使是一种蛋白的不同亚基也将涉及多个基因的协调表达。
- 真核生物DNA在细胞核内与多种蛋白质结合构成染色质,这种复杂的结构直接影响着基因表达。
真核生物的遗传信息不仅存在于核DNA上,还存在线粒体DNA上,核内基因与线粒体基因的表达调控既相互独立而又需要协调。
真核与原核在调控机制上的主要差别
调控的原因
- 原核生物基因表达调节的目的是为了更有效的更经济地对环境的变化做出反应
- 真核生物基因表达调节的主要目的是细胞分化,有时空特异性
- 调控的层次
- 原核生物基因表达调控主要集中在转录水平
- 真核生物,转录后水平核转录水平各占“半壁江山”,而某些调控层次是真核生物特有的,比如染色质水平,RNA后加工水平和mRNA运输等
调控的手段
活性染色质(active chromatin):当基因被激活时,染色质相应区域发生某些结构与性质变化,这些具有转录活性的染色质被称为活性染色质。
- 转录活化的染色质对核酸酶极为敏感
- 转录活化染色质的组蛋白发生变化
- CpG岛甲基化水平降低
- DNA甲基化:真核基因组中胞嘧啶的第5位碳原子可以在DNA甲基转移酶(DNA methyltransferase)的作用下被甲基化修饰为5-甲基胞嘧啶,并且以序列CG中的胞嘧啶(C)甲基化更加常见。
- CpG岛(CpG island):真核基因组中GC含量达60%,长度为300~3000bp的区段称作CpG岛。CpG岛主要位于基因的启动子和第一外显子区域,约有60%以上基因的启动子含有CpG岛
- CpG岛甲基化对基因表达的影响:CpG岛的高甲基化促进染色质形成致密结构,不利于基因表达;CpG岛的低甲基化作用正好相反。
- 表观遗传:甲基化修饰可以发生遗传,DNA甲基化酶可以对子链DNA相同位置进行甲基化
转录起始的调节
顺式作用元件是转录起始的关键调节部位
顺式作用元件:是指可影响自身基因表达活性的DNA序列,通常是非编码序列,但是并非都位于转录起始点上游。真核基因的顺式作用元件分为启动子、增强子及沉默子等。
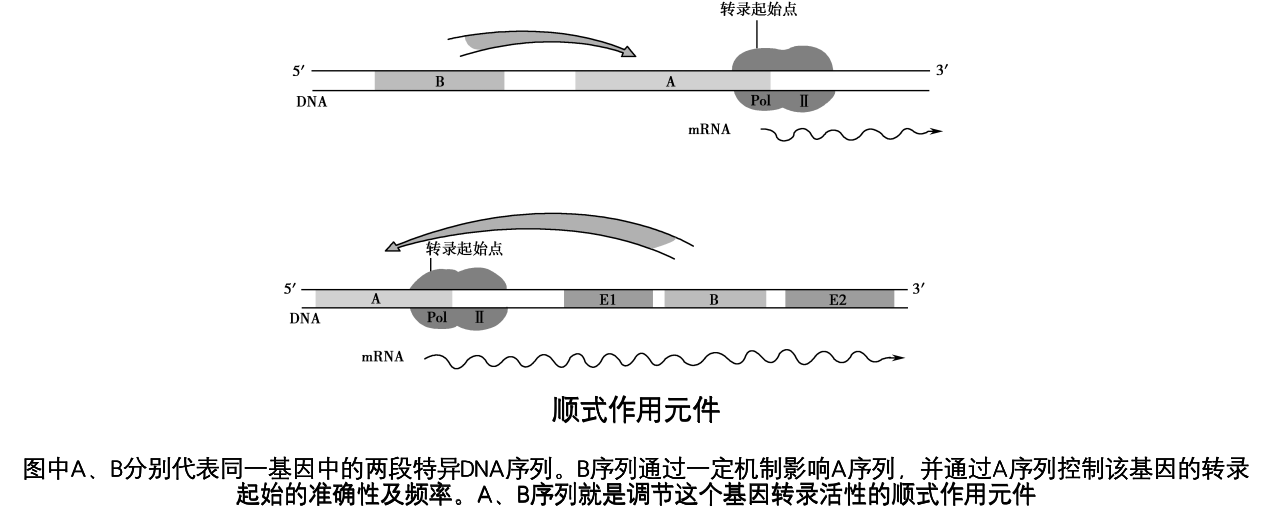
真核生物启动子结构和调节远较原核生物复杂
- 启动子(promoter):一般位于转录起始点上游,约为100~200bp序列,包含有若干具有独立功能的DNA序列元件,每个元件约长7~30bp。
典型的Ⅱ类启动子由TATA盒或下游启动子元件(downstream promoter element,DPE)和起始元件(initiator element,Inr)以及上游调控元件组成。
增强子是一种能够提高转录效率的顺式调控元件
增强子(enhancer):长度大约是200bp,可使旁侧的基因转录效率提高100倍或更多。
增强子的作用
沉默子(silencer):是一类基因表达的负性调控元件,当其结合特异蛋白因子时,对基因转录起阻遏作用。
沉默子的作用:与增强子类似,其作用亦不受序列方向的影响,也能远距离发挥作用,并可对异源基因的表达起作用。
绝缘子阻碍其他调控元件的作用
绝缘子(insulator):位于增强子或沉默子与启动子之间,与特异蛋白质因子结合后,阻碍增强子或沉默子对启动子的作用。
绝缘子还可位于常染色质与异染色质之间,保护常染色质的基因表达不受异染色质结构的影响;与增强子类似,发挥作用与序列的方向性无关。
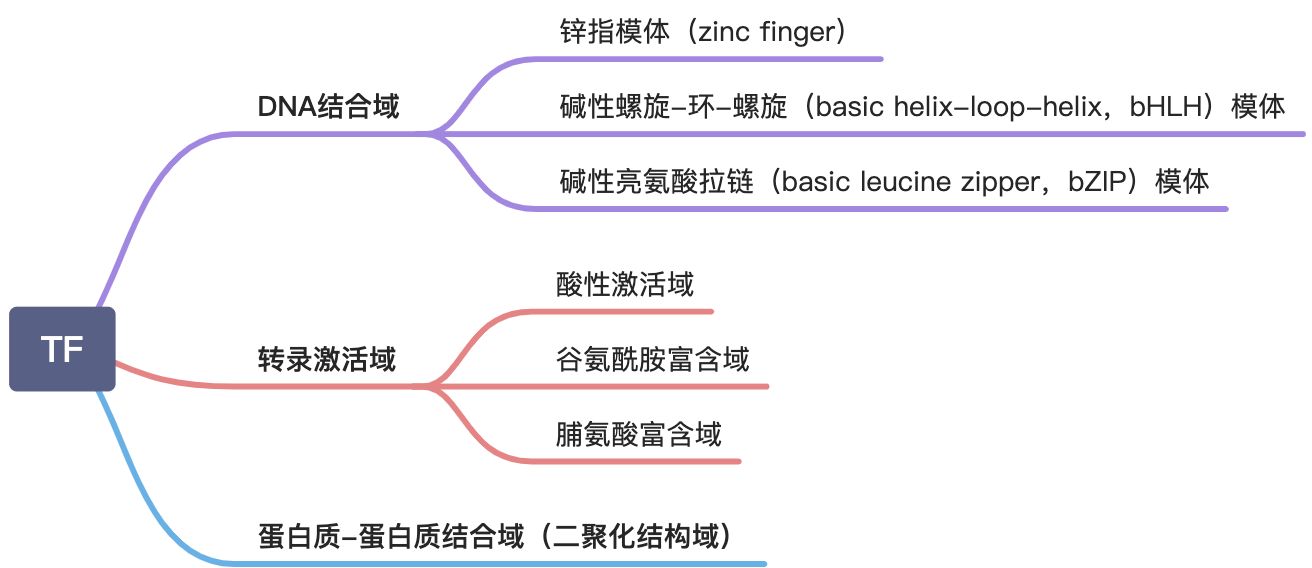
转录因子是转录起始调控的关键分子
转录因子(transcription factor):是指真核基因的转录调节蛋白,由其编码基因表达后,进入细胞核,通过识别、结合特异的顺式作用元件而增强或降低相应基因的表达。转录因子也被称为反式作用蛋白或反式作用因子。
- 顺式作用蛋白:有些基因产物可特异识别、结合自身基因的调节序列,调节自身基因的开启或关闭,这就是顺式调节作用。具有这种调节方式的调节蛋白称为顺式作用蛋白。
转录因子分类
RNA聚合酶介导基因转录时所必需的一类辅助蛋白质,帮助聚合酶与启动子结合并起始转录,对所有基因都是必需的。例如: TFⅡD
-
特异转录因子
为个别基因转录所必需,决定该基因表达的时间空间特异性。
- 包括转录激活因子和转录抑制因子。
- 受环境影响,是使环境变化在基因表达水平得到体现的关键分子。
-
转录因子的结构特点
转录起始复合物的组装是转录调控的主要方式
真核生物主要有三种RNA聚合酶,分别负责催化生成不同的RNA分子。其中RNA聚合酶Ⅱ参与转录生成所有mRNA前体及大部分snRNA
-
转录后调控主要影响真核mRNA的结构与功能
mRNA的稳定性影响真核生物基因表达
5′-端的帽结构可以增加mRNA的稳定性
使得mRNA免于在5′-核酸外切酶的作用下被降解,从而延长了mRNA的半衰期
通过与相应的帽结合蛋白结合而提高翻译的效率,并参与mRNA从细胞核向细胞质的转运
3′-端的poly(A)尾结构防止mRNA降解
Poly(A)及其结合蛋白可以防止3′-核酸外切酶降解mRNA,增加mRNA的稳定性。
-
一些非编码小分子RNA可引起转录后基因沉默
核酶、细胞核小分子RNA(snRNA),核仁小分子RNA(snoRNA),miRNA以及siRNA,都可引起转录后基因沉默
mRNA前体的选择性剪接可以调节真核生物基因表达
mRNA前体的选择性剪接使一条mRNA前体产生了不同的成熟mRNA,并由此产生了完全不同的蛋白质,显示了基因调控对生物多样性的决定作用。
抗体基因多样性产生的分子机制
VJ和VDJ的重组连接
- 连接的多样性
- 插入的多样性,DJ连接或 VDJ连接中随机插入若干个核苷酸
- 体细胞超突变,主要发生在V基因
-
真核基因表达在翻译及翻译后仍可受到调控
对翻译起始因子活性的调节主要通过磷酸化修饰进行
翻译起始因子eIF-2α的磷酸化抑制翻译起始
-
翻译起始因子eIF-2α的磷酸化抑制翻译起始
蛋白质合成速率的快速变化在很大程度上取决于起始水平,通过磷酸化调节翻译起始因子(eukaryotic initiation factor, eIF)的活性对起始阶段有重要的控制作用。
eIF-4E及eIF-4E结合蛋白的磷酸化激活翻译起始
帽结合蛋白eIF-4E与mRNA帽结构的结合是翻译起始的限速步骤,磷酸化修饰及与抑制物蛋白的结合均可调节eIF-4E的活性。
磷酸化的eIF-4E,与帽结构的结合力升高了4倍,提高翻译的效率。
RNA结合蛋白参与了对翻译起始的调节
RNA结合蛋白(RNA binding protein, RBP),是指那些能够与RNA特异序列结合的蛋白质。
- 基因表达的许多调节环节都有RBP的参与,如前述转录终止、RNA剪接、RNA转运、RNA胞浆内稳定性控制以及翻译起始等。
如IRE结合蛋白(IRE-BP)调节铁转运蛋白受体mRNA稳定性。铁浓度低,IRE-BP活化结合IRE,阻碍核糖体小亚基与mRNA结合,抑制翻译起始。
翻译后调控:对翻译产物修饰或降解速度调控
新合成蛋白质的半衰期长短是决定蛋白质生物学功能的重要影响因素。因此,通过对新生肽链的水解和运输,可以控制蛋白质的浓度在特定的部位或亚细胞器保持在合适的水平。
许多蛋白质需要在合成后经过特定的修饰才具有功能活性。通过对蛋白质的可逆的磷酸化、甲基化、酰基化修饰,可以达到调节蛋白质功能的作用,是基因表达的快速调节方式。
小分子RNA对基因表达的调节
微小RNA (microRNA, miRNA)
是一大家族小分子非编码单链RNA,长度约20-25个碱基,由一段具有发夹环结构,长度为70~90个碱基的单链RNA 前体(pre-miRNA)经Dicer酶剪切后形成。
- 这些成熟的miRNA 与其他蛋白质一起组成RNA诱导的沉默复合体(RNA-induced silencing complex, RISC),通过与其靶mRNA 分子的3’ 端非翻译区域(3’ UTR)互补匹配,再以目前尚不清楚的机制抑制该mRNA 分子的翻译。
miRNAs的特点
- miRNAs是一种20-25nt的单链RNA, 广泛存在于真核生物中,不编码蛋白质,它本身不具有开放阅读框(ORF)
- 成熟的miRNAs,是由具有发夹结构的约70-90个碱基大小的单链RNA前体经过Dicer酶加工后生成
- miRNA基因常以单拷贝、多拷贝或基因簇等多种形式存在于基因中。70%~90% 位于基因间隔区,其余在内含子,极少数在蛋白编码区。
- miRNA不仅在结构上保守,而且在物种间具有高度的进化保守。(5’端的2-8个核苷酸有一定保守性)
- miRNA具有明显的时空特异性及组织特异性。lin-4与lin-7呈时间特异性表达;miR-1 在肌肉组织中特异表达,并且调控肌肉组织的生长、分化。
小干扰RNA(small interfering RNA, siRNA)
是细胞内一类双链RNA(dsRNA),在特定情况下通过酶切机制,转变为具有特定长度(21-23个碱基)和特定序列的小片段RNA。
- 双链siRNA参与RISC组成,与特异的靶mRNA完全互补结合,导致靶mRNA降解,阻断翻译过程。
- 由siRNA介导的基因表达抑制作用被称为RNA干涉(RNA interference,RNAi)。
- RNAi机制
- 在起始步骤,加入的dsRNA被Dicer酶切割为21-23核苷酸长的小分子干扰RNA片段
- 在RNAi效应阶段,siRNA双链结合一个核酶复合物从而形成所谓RNA诱导沉默复合物(RNA-induced silencing complex, or RISC)
- 激活RISC需要一个ATP依赖的将siRNA解双链的过程
- 激活的RISC通过碱基配对定位到同源mRNA转录本上,并在距离siRNA3’端12个碱基的位置切割mRNA
- RNA干扰指导的基因沉默的结果
- Degradation of the target mRNA (引起靶标mRNA的降解)
- Inhibition of translation of the target mRNA (抑制靶标mRNA的翻译)
- Silencing the gene transcription from the target promoter (引起靶标启动子的转录沉默).
siRNA和miRNA的差异比较
|
| siRNA | miRNA | | :—-: | :—-: | :—-: | | 前体 | 内源或外源长双链RNA诱导产生 | 内源发夹环结构的转录产物 | | 结构 | 双链分子 | 单链分子 | | 功能 | 降解mRNA | 阻遏其翻译 | | 靶mRNA 结合 | 需完全互补 | 不需完全互补 | | 生物学效应 | 抑制转座子活性和病毒感染 | 发育过程的调节 |
IncRNA在基因表达中的作用
- 长链非编码RNA(lncRNA)是一类转录本长度超过200个核苷酸的RNA分子,不直接参与基因编码和蛋白质合成,但是可在表观遗传水平、转录水平和转录后水平调控基因的表达。

若有收获,就点个赞吧
0 人点赞
