
责编 | 酶美
DNA 胞嘧啶甲基化是一种可逆的表观遗传修饰,参与调控基因表达和染色质开放程度,广泛影响个体发育、癌症发生和衰老等生物学过程。哺乳动物甲基化组 (methylome) 由两个相互拮抗的生物学作用共同形成,即 DNA 甲基转移酶 DNMT 介导的甲基化 (methylation) 和 TET 双加氧酶介导的去甲基化 (demethylation)。这两个酶家族在 DNA 上的结合呈现竞争模式【1】 ,敲降Dnmt3a后 TET1 蛋白在基因启动子区域(promoter)的结合增强,反之亦然【2】 。前人研究表明,Dnmt3a & Tet2 双敲除小鼠的存活率低于Dnmt3a或Tet2单敲除小鼠【3】 。并且,在人类 T 细胞淋巴瘤中, DNMT3A 和 TET2 的突变呈现显著的并存【4】 。这些发现表明甲基化和去甲基化过程的并发与肿瘤发生密切相关。然而,目前没有方法能够衡量两者的并发,其与基因表达的关联也未被揭示。
近日,来自美国加州大学尔湾分校李蔚教授团队的史偈君等研究人员在Nature Communications杂志发表文章The concurrence of DNA methylation and demethylation is associated with transcription regulation,开发了基于亚硫酸氢盐测序 (bisulfite-seq) 的计算生物学方法——甲基化并发率 (methylation concurrence ratio),能够衡量甲基化和去甲基化过程在基因调控元件上的拮抗作用,具有不同于传统平均甲基化 (average methylation ratio) 的生物学特性,并可作为抑癌基因的新型表观遗传分子标记。

自甲基化测序方法面世以来,DNA 甲基化水平一直以 “平均” 的方式进行量化,但这种方法无法刻画甲基化的局部变异。
如图 1a 所示,虽然两个例子的甲基化模式存在明显差异,但平均甲基化却都是 0.5。Example 2 中存在较多的不完全甲基化片段,在这些片段上可能同时存在甲基化与去甲基化作用。因此,研究人员将不完全甲基化片段中的未甲基化 CpG 定义为“甲基化并发 CpG”(图 1b 中红色所示),甲基化并发率即为并发 CpG 的比率。
如图 1b 所示,与平均甲基化相比,甲基化并发率对甲基化模式的局部变异更加敏感。
如图 1c和d 所示,研究人员分析了 DNMT 和 TET 酶的 ChIP-seq 数据,并定义了基因启动子区的 “DNMT-TET 共结合指数”。该指标越大,表明 DNMT 和 TET 酶在启动子区的共结合效应越强;反之,若该指标为 0,则表明两者不存在共结合。该共结合指数与甲基化并发率之间存在显著的正相关性(Spearman’s R = 0.47,图 1d),而与平均甲基化的相关性并不显著(图 1c)。
这表明甲基化并发率可以刻画 DNMT 和 TET 酶的共结合,而平均甲基化则不能。
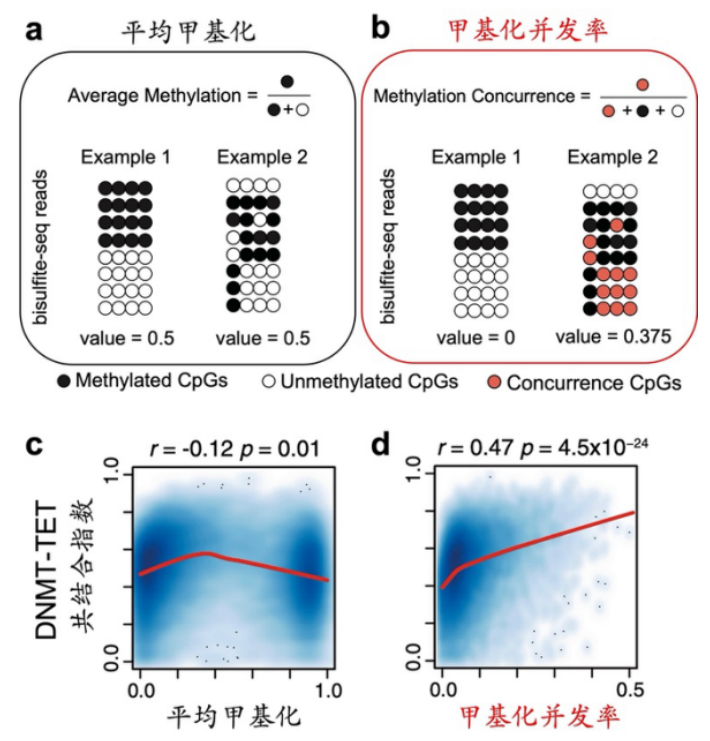
图 1. 平均甲基化与甲基化并发率方法的比较
虽然平均甲基化方法已被广泛使用,但其与基因表达的关系仍不完全明确。目前普遍认为启动子区 CpG 岛(CpG island)的平均甲基化与基因表达水平呈负相关,但是基因体区域(gene-body)和增强子区域(enhancer)的平均甲基化与基因表达的关系仍存在争议。本研究发现,不同类型调控元件的平均甲基化与基因表达水平的相关性不尽相同。而甲基化并发率能够更好的指示基因表达水平,不论哪一种基因调控元件(例如启动子、基因体和增强子)上的甲基化并发率始终与基因表达呈现较强的负相关性。之后,研究人员发现抑癌基因在正常样本中的甲基化并发率水平很低,并且该现象在不同组织中高度保守。通过分析 TCGA 中多种癌症(子宫肌瘤、乳腺癌等)样本的甲基化组,进而发现肿瘤中甲基化并发率的升高与 40~60% 的抑癌基因表达下调有关,而这些基因的平均甲基化却未发生改变。总之,本研究提出了甲基化并发率这一新型甲基化量度方法,强调了甲基化局部变化对基因表达的显著影响,拓展了我们对肿瘤异常甲基化模式的理解。
文章的第一作者为史偈君博士,通讯作者为李蔚教授。
文章原文链接:https://www.nature.com/articles/s41467-021-25521-7
公众号原文链接:https://mp.weixin.qq.com/s/-GwzQQabEHGxkMXQatiRcQ
Code availability:https://github.com/JiejunShi/CAMDA,Concurrence of Active Methylation and De-methylAtion (CAMDA)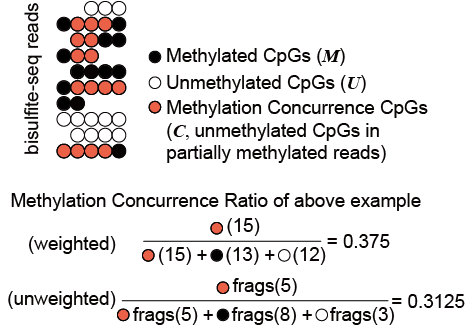
图 1. 亚硫酸氢盐测序捕获的甲基化并发示意图。
亚硫酸氢盐测序reads分为三类片段,即甲基化(M)片段(图1中连续的实心圆圈)、未甲基化(U)片段(连续的空白圆圈)和甲基化-并发(C)片段(连续的红色圆圈) .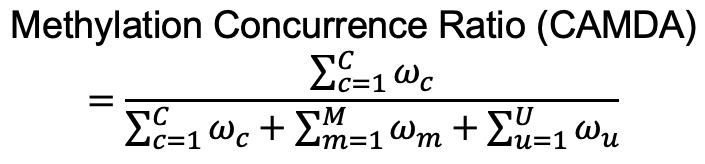
图 2. 甲基化并发率的定义。
基因组区域的甲基化并发率定义为C片段权重的总和除以该区域中所有片段的权重总和。每个片段的权重可以设置为其 CpG 的数量(在下面的第1节中)或 1(未加权,在下面的第2节中)。’ M ‘、’ U ‘和’ C ‘分别代表甲基化片段、未甲基化片段和甲基化同时片段的数目。’ω m ‘、’ω u ‘ 和’ω c ‘ 是每个片段的权重。
参考文献
- Charlton, J. et al. TETs compete with DNMT3 activity in pluripotent cells at thousands of methylated somatic enhancers. Nat Genet 1–9 (2020) doi:10.1038/s41588-020-0639-9.
- Gu, T. et al. DNMT3A and TET1 cooperate to regulate promoter epigenetic landscapes in mouse embryonic stem cells. Genome Biol 19, 88 (2018).
- Zhang, X. et al. DNMT3A and TET2 compete and cooperate to repress lineage-specific transcription factors in hematopoietic stem cells. Nat Genet 48, 1014 1023 (2016).
- Couronné, L., Bastard, C. & Bernard, O. A. TET2 and DNMT3A mutations in human T-cell lymphoma. _New Engl J Medicine_366, 95–6 (2012).
[
](https://mp.weixin.qq.com/s/-GwzQQabEHGxkMXQatiRcQ)
思考:这种以片段/区域的甲基化水平当成基因组甲基化水平的基本单位,之前也有人报道和研究过,类似研究的出发点也是这篇文章的核心:“平均” 的方式进行量化基因组甲基化水平,这种方法无法刻画甲基化的局部变异。所以可以尝试这种方法,但是后续的分析步骤也可能随之改变。
综述-单细胞DNA甲基化研究基础篇:从实验策略到数据分析方法简介

若有收获,就点个赞吧
0 人点赞
