导读:10 年前,我们开启了细胞核内 miRNA 的研究;5 年前,我们发表了第一项核内 miRNA 研究成果,发现并定义了一类在细胞核内具有激活功能的 miRNA——NamiRNA (Nuclear activating miRNA),提出了 NamiRNA - 增强子 - 基因激活的新理论;5 年后的今天,我们发表了 NamiRNA 第二项重要研究成果,揭示了肿瘤中抑癌基因沉默的新机制,为唤醒沉默的抑癌基因提供了全新的思路。
抑癌基因失活是细胞癌变过程中常发生的事件。DNA 甲基化可调控基因的活性,启动子区域的高甲基化通常会抑制基因表达,这可以解释细胞癌变过程中部分抑癌基因的失活。然而,大量的抑癌基因启动子区域并没有出现所预想的高甲基化,这些抑癌基因在肿瘤中的低表达就成了肿瘤领域的未解之谜,如何唤醒沉睡的抑癌基因更成了肿瘤研究领域可望而不可及的梦想。
癌基因和抑癌基因是我们生物医学领域耳熟能详的名词,癌基因的高表达或获得性突变以及抑癌基因的低表达或缺失性突变被认为是肿瘤发生的最重要的原因 (1,2)。抑癌基因一个显著的特征是在肿瘤中低表达。我们不禁要问两个重要的问题:1)抑癌基因为什么会在肿瘤中的低表达?2)如何激活抑癌基因用于肿瘤的治疗?
关于抑癌基因为何在肿瘤中低表达,这涉及肿瘤表观遗传学领域早期的著名争论——肿瘤到底是与低甲基化还是高甲基化有关,这一争论时间跨度长达 15 年。这就不得不提到肿瘤甲基化领域的 “三驾马车”,他们共同推进了肿瘤 DNA 甲基化领域的研究。第一位就是我在美国博士后期间的老板、约翰 · 霍普金斯大学 Andrew Feinberg 教授,他于 1983 年在 Nature 杂志发文揭示了全基因组水平的低甲基化是肿瘤的重要特征,他是将 DNA 甲基化与肿瘤联系起来的第一人。但有意思的是,他在这篇文章中说不清肿瘤的低甲基化是如何导致肿瘤的。两年后的 1985 年,约翰 · 霍普金斯大学另一位著名的教授 Stephen Baylin 发现抑癌基因的启动子区域存在高甲基化,进而阐明启动子区域高甲基化导致抑癌基因的低表达是肿瘤发生的重要原因。这就形成了一个悖论,肿瘤的发生到底是因为 DNA 低甲基化还是高甲基化,一时间 DNA 高甲基化占据上风,因为启动子区域的高甲基化导致抑癌基因的下调很好的符合了肿瘤发生的 “二次打击” 学说。对这件事起到推波助澜作用的还有另一位肿瘤 DNA 甲基化领域的著名教授 Peter Jones,他于 1977 年发现核酸类似物 5-azacytidine (5-AzaC) 可以显著上调基因的表达,最后发现 5-Aza-C 主要与 DNA 去甲基化有关,大约在 1990 年前后基本上奠定了抑癌基因的高甲基化与肿瘤发生的稳定关系,这逐渐形成了肿瘤领域的共识,推动了 DNA 去甲基化药物用于肿瘤治疗。
尽管 DNA 高甲基化导致抑癌基因的下调已成为肿瘤领域的共识,但并非无懈可击:1)抑癌基因有接近 1000 多个,但常研究的抑癌基因大约 50 个,那么大部分抑癌基因是怎样下调的呢?启动子甲基化解释所有抑癌基因的下调显然力不从心。2)5-AzaC 是一个广谱的 DNA 甲基化抑制剂,对抑癌基因的激活没有选择性,其肿瘤治疗效果有限,如何激活抑癌基因就成了肿瘤领域的一个 “烫手山芋”,人人想吃,但无从下口。这就催生了一个重要的问题:启动子甲基化导致抑癌基因下调是普遍规律吗?是否还有其他机制?是否能够基于这种机制,特异性地激活被抑制的抑癌基因,为肿瘤治疗带来新的曙光?
复旦大学于文强课题组自 2009 年起致力于核内 miRNA 激活研究,提出 NamiRNA (Nuclear activating miRNA) 通过增强子激活基因表达的全新理论。近日,复旦大学于文强课题组联合北京中科院生态中心汪海林课题组以及复旦大学附属肿瘤医院李大强课题组,揭示了 NamiRNA - 增强子介导的抑癌基因的失活机制,并发现可通过 NamiRNA - 增强子网络可重新激活抑癌基因,提示 NamiRNA - 增强子可作为癌症治疗的全新靶点。相关研究于 2021 年 7 月 30 日在线发表在 Nucleic Acid Research 杂志(下文简称 NAR)杂志(图 1)。

图 1. 相关工作发表在 2021 年 7 月 30 日的 NAR
启动子高甲基化失活抑癌基因?
Not the Whole Picture!
DNA 甲基化能改变染色质结构和 DNA 构象与稳定性,启动子的 DNA 甲基化可以导致转录沉默,进而引起基因的低表达或者失活 (3),如常见的抑癌基因 VHL (Von Hippel Lindau 肿瘤抑制因子),p16INK4a (细胞周期蛋白依赖性激酶抑制剂 2A[CDKN2A]),BRCA1 (乳腺癌易感性基因 1) 和 hMLH1 (人类错配修复基因) 等在肿瘤中低表达是由于启动子区域高甲基化 (4,5)。然而,对于 TSGene 数据库公布的 983 个肿瘤中低表达的抑癌基因 (6,7),抑癌基因低表达与启动子高甲基化关联的报道并不多。那么,其中有多少个基因的低表达是抑癌基因启动子甲基化引起的?抑癌基因在肿瘤中表达下调是否还有其他因素?本次研究通过系统的生信分析发现,乳腺癌中有 622 个抑癌基因,其中 383 个抑癌基因低表达。出乎意料的是,与正常对照组相比,乳腺癌中约 68% 低表达的抑癌基因启动子甲基化状态没有发生显著高甲基化(图 2)。

图 2. 乳腺癌中低表达抑癌基因的启动子甲基化分析。秩和检验分析结果表明,和正常对照相比,乳腺癌中 68.2% 的低表达抑癌基因启动子没有高甲基化(红色框标注)。
NamiRNA - 增强子 - 基因激活通路
The Neglected but Important Player!
既然在乳腺癌中启动子高甲基化无法解释抑癌基因的低表达现象,那么抑癌基因的表达受到哪些因素调控呢?鉴于增强子与启动子一样是顺式调控元件,本研究选取这 68% 没有甲基化差异的抑癌基因,对它们的增强子标记 H3K27ac 在乳腺癌样本和正常对照中的富集进行了分析,结果显示有相当一部分抑癌基因(45/212)的增强子上 H3K27ac 在乳腺癌组织中的富集程度要显著低于正常对照组,提示增强子可能与乳腺癌中抑癌基因低表达相关。
miRNA 作为一种长度为 18~23 个核苷酸的非编码小 RNA,通常被认为在细胞质中通过结合靶基因的 3’-UTR 降解 mRNA 或抑制翻译从而发挥负向调控作用。随着 miRNA 研究的不断深入和深度测序技术的发展,越来越多的证据表明,miRNA 不仅可定位于细胞质,而且也存在于其他细胞器中,miRNA 的不同细胞定位影响 miRNA 的功能 (8)。
复旦大学于文强课题组专注于细胞核内 miRNA 研究十余年,在前期工作中对 ENCODE 数据库中 7 种不同细胞系 1594 条 miRNA 前体进行系统分析,发现 300 多条 miRNA 前体在基因组中的位置与增强子高度重叠,这些 miRNA 包括 hsa-miR-24、hsa-miR-3179 等;miR-3179 被证实能激活邻位基因 ABCC6 和 PKD1P1,miR-24-1 能激活靶基因 FBP1 和 FANCC 的表达 (9)。这类 miRNA 定位于细胞核中并且通过靶向增强子进而激活临近或远端基因,这类细胞核内具有激活功能的 miRNA 定义为 NamiRNA (Nuclearactivating RNA)(9,10)。相关研究于 2017 年发表在 RNA Biology,Google Scholar 显示该文章至今被引 154 次。

图 3. NamiRNA 相关工作的原始文章
在肿瘤中,大量研究已经揭示了 miRNA 低表达与肿瘤发生发展的关联。miRNA 在肿瘤中低表达是肿瘤的另一个重要特征,很多 miRNA 在正常组织中的表达要高于肿瘤 (11)。如 let-7 家族 miRNA 可负向调控 MYC 及 RAS 家族癌基因的表达从而抑制肿瘤发展,且 let-7 家族 miRNA 在多种癌症中低表达并与不良预后相关 (12,13);肝脏细胞中 miR-199a 和 miR-199b 表达下调,与肝癌的发生发展有关 (14)。另一方面,增强子是一类促进基因转录的 DNA 顺式调控元件,可近距离或远距离调控基因的表达,在细胞分化、组织发育、个体生长和肿瘤发生发展过程中发挥重要作用。H3K27ac 是激活状态增强子 (active enhancer) 的标记。增强子相关的染色质修饰异常引起抑癌基因的增强子失活或者癌基因增强子的异常激活都与肿瘤的发生有密切关系 (15)。因此,本研究将 miRNA 和增强子两个重要的调控元件联系起来。
miR-339 - 增强子 -GPER1 调控
Too Good to Be True
乳腺癌是全球发病率最高的癌症,也位列我国女性恶性肿瘤发病首位。三阴性乳腺癌 (Triple-negative breast cancer, TNBC) 具有高侵袭性、易转移的特点,目前尚无有效治疗手段,因此寻找新的靶点和有效治疗药物迫在眉睫。
结合 NamiRNA - 增强子 - 基因激活通路的研究 (9) 以及乳腺癌中增强子的异常,我们推测乳腺癌中可能存在大量低表达的 NamiRNA,进而通过 NamiRNA - 增强子通路,丧失了对抑癌基因的激活作用,最终导致了抑癌基因的低表达。
首先我们分析得出乳腺癌中低表达的 NamiRNA 候选基因,通过 mRNA-seq 分析(图 4)以及绝对定量 PCR 实验验证,优先筛选出在乳腺癌中低表达的 miR-339 和靶向激活的抑癌基因 GPER1 作为研究对象。

图 4. mRNA-seq 分析在乳腺癌细胞 4175 中过表达 miR-339 激活的基因,其中包含GPER1
有意思的是,GPER1 是同一条染色体(Chr7)上距离 miR-339 最近的抑癌基因,在 UCSC Genome Browser 里可以查到 miR-339 就在 GPER1上游 60kb 的位置,并且 ENCODE 数据也显示 miR-339 的基因座位置的 H3K27ac 富集(图 5)。

图 5. UCSC 上 miR-339 和 GPER1 在 7 号染色体上的位置。UCSC 截图显示 ENCODE 数据中 miR-339 位置有显著的 H3K27ac 富集,提示 miR-339 和增强子位置重叠,其中不同的颜色代表不同的细胞类型。
为了进一步验证,本研究收集了 157 对不同分子分型的乳腺癌组织(Luminal A,Luminal B,HER2 富集型和三阴性乳腺癌)及相应的癌旁对照组织进行了 RT-qPCR 检测,结果表明与对照组织相比,miR-339 和抑癌基因 GPER1 在 Luminal 型及三阴性乳腺癌组织中表达下调,而在 HER2 富集型乳腺癌组织中没有明显差异(图 6)。同时,miR-339 可作为乳腺癌潜在的分类标记,对于 miR-339 低表达的乳腺癌患者,通过 miR-339 再激活 GPER1,有可能带来福音。

图 6. 不同类型乳腺癌组织中 miR-339 和 GPER1 表达情况和相关关系
那么,miR-339 和抑癌基因 GPER1 对乳腺癌细胞行为学会产生怎样的影响呢?首先,在 Luminal 乳腺癌细胞系 T47D 和三阴性乳腺癌细胞系 4175 中过表达外源性 miR-339,结果提示在这两个非 HER2 富集型乳腺癌细胞中过表达 miR-339 后抑癌基因 GPER1 的表达均被激活,并且它们的增殖和克隆形成能力均受到明显抑制,表明过表达 miR-339 可再激活(reactivate)抑癌基因 GPER1,进而抑制乳腺癌细胞生长及增殖;也提示 miR-339 可以作为三阴性乳腺癌的治疗提供潜在的靶点(图 7)。
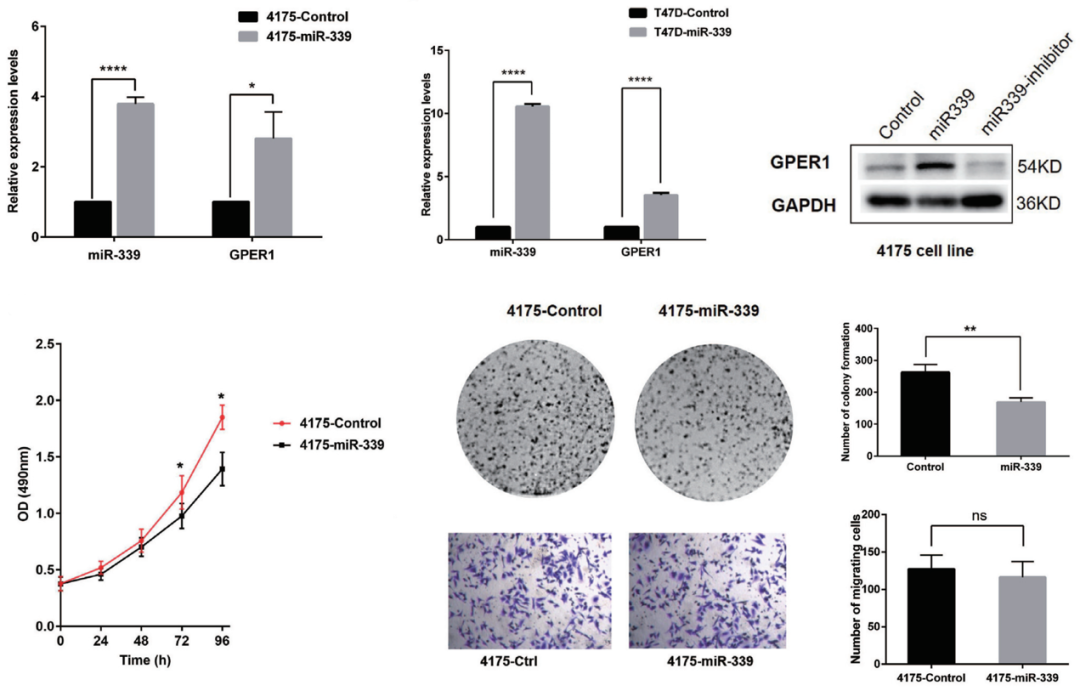
图 7. miR-339 过表达可重新激活抑癌基因 GPER1 并抑制乳腺癌细胞生长和增殖
此外,机制研究上,本研究通过双荧光素报告基因实验证明了 miR-339 locus 具有增强子活性;ChIP-seq 结果表明过表达 miR-339 可以提高靶基因增强子活性(图 8);另一方面,利用 CRISPR/Cas9 敲除技术破坏增强子序列完整性后,miR-339 不能再激活 GPER1(图 9)。增强子就像开关一样,过表达 NamiRNA 会提高增强子活性,打开促进靶基因表达的开关;反之,NamiRNA 低表达就会降低增强子活性,关闭促进靶基因表达的开关。

图 8. ChIP 高通量测序分析。过表达 miR-339 可以激活增强子活性,ChIP-seq 分析过表达 miR-339 后,miR-339 DNA locus 位置附近 H3K27ac 富集明显高于对照组。
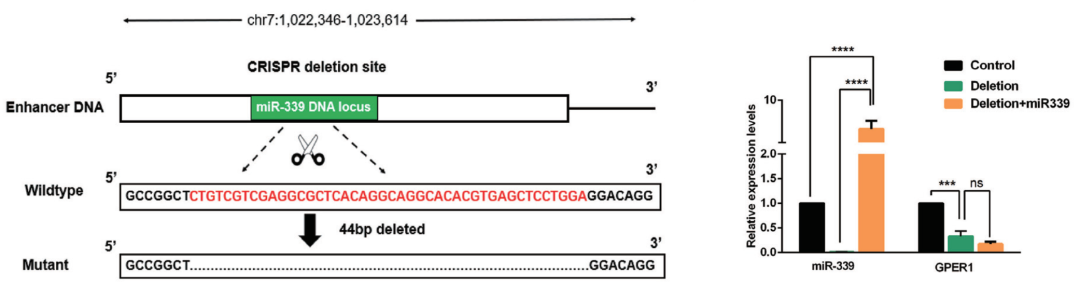
图 9. CRISPR 系统破坏增强子完整性后,过表达 miR-339 不能激活 GPER1
NamiRNA 激活沉默的抑癌基因潜力无限
Too Simple but Powerful!
为了探究过表达 miR-339 再激活 GPER1 是否可以对乳腺癌细胞发挥抑制作用,我们进行了动物荷瘤实验。结果表明,过表达 miR-339 可以抑制乳腺癌发展与增殖,提示 miR-339 是乳腺癌潜在的治疗靶点,可为乳腺癌治疗提供新的线索和思路(图 10)。

图 10. 动物荷瘤实验。NamiRNA 作用不仅有效而且特异性,提示 NamiRNA 做为小核酸药物的巨大潜力
NamiRNA 不怕做不到,就怕想不到
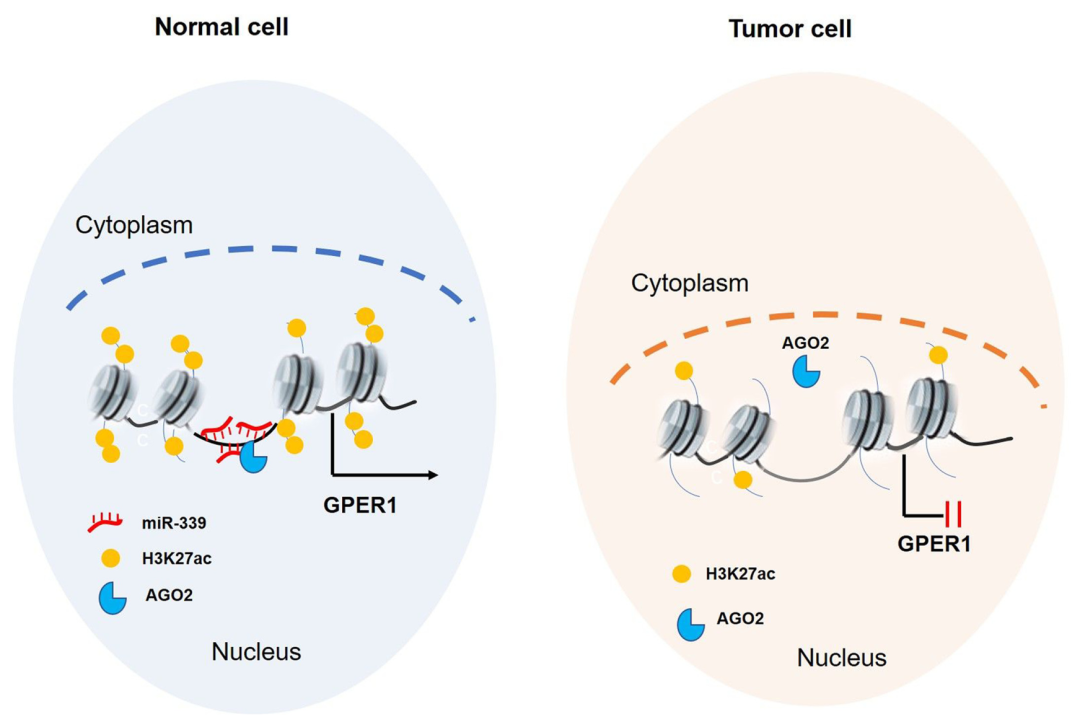
图 11. 在正常和肿瘤细胞中 miR-339 - 增强子介导的抑癌基因 GPER1 的活性
本研究阐明了抑癌基因在肿瘤中的低表达与增强子及 miRNA 的异常调控有关。本次工作在分子机制上对 miR-339 通过增强子靶向激活抑癌基因 GPER1 进行了较为详细的阐述。此外,乳腺癌中抑癌基因 GPER1 低表达与 miR-339 低表达有关,细胞行为学实验及动物实验结果均表明过表达 miR-339 后可再激活抑癌基因 GPER1 进而抑制乳腺癌细胞的生长与增殖,提示 miR-339 可为乳腺癌尤其是三阴性乳腺癌提供潜在的治疗靶点和策略。
后记
NamiRNA 研究开展 10 年多,由最初的假说到实验验证,实验室的学生走了一波又一波,套用我们实验室学生的话说,在 NamiRNA 研究的星辰大海中,谁信不重要,自己信最重要。NamiRNA 研究已经成为我们实验室重要的研究方向,先后有 10 余人投入其中,涉及 NamiRNA 作用机制、重要表观遗传现象的另类解释、疾病治疗新策略、重要传染性疾病新的预防方案等。我们深知一个全新的研究方向要得到科学共同体的认可需要时间,对科学研究者来说,我们坚信科学真理不是用来膜拜的,而是用来颠覆和超越的,只要方向对了,到达胜利的终点只是时间问题。这世界本没有路,总得有人去披荆斩棘。感谢汪海林老师、李大强老师,相信本身就是一种无穷的力量。昨天我们孤独前行,今天我们结伴而行,明天我们将一起大步前行!
撰文: 梁英、徐鹏、于文强。文中 “我” 为于文强教授。
参考文献:
- Macleod, K. (2000) Tumor suppressor genes. Curr Opin Genet Dev, 10, 81-93.
- Sherr, C.J. (2004)Principles of tumor suppression. Cell,116, 235-246.
- Skvortsova, K., Stirzaker, C. and Taberlay, P. (2019) The DNA methylation landscape in cancer. Essays Biochem, 63, 797-811.
- Esteller and Manel. (2002) CpG island hypermethylation and tumor suppressor genes: a booming present, a brighter future. Oncogene,21, 5427-5440.
- Esteller, M.(2007) Epigenetic gene silencing in cancer: the DNA hypermethylome. Human Mol Genet, 16 Spec No 1, R50-59.
- Zhao, M., Kim, P., Mitra, R., Zhao, J. and Zhao, Z. (2016) TSGene 2.0: an updated literature-based knowledgebase for tumor suppressor genes. Nucleic Acids Res, 44, D1023-1031.
- Zhao, M., Sun, J.and Zhao, Z. (2013) TSGene: a web resource for tumor suppressor genes. Nucleic Acids Res, 41, D970-976.
- Thurman, R.E., Rynes, E., Humbert, R., Vierstra, J., Maurano, M.T., Haugen, E., Sheffield, N.C., Stergachis, A.B., Wang, H., Vernot, B. et al. (2012) The accessible chromatin landscape of the human genome. Nature, 489, 75-82.
- Xiao, M., Li, J., Li, W., Wang, Y., Wu, F.Z., Xi, Y.P., Zhang, L., Ding, C., Luo, H.B., Li, Y. et al. (2017) MicroRNAs activate gene transcription epigenetically as an enhancer trigger. RNA Biol, 14, 1326-1334.
- Liang, Y., Zou, Q. and Yu, W. (2017) Steering Against Wind: A New Network of NamiRNAs and Enhancers. Genomics Proteomics Bioinformatics, 15, 331-337.
- Lu, J., Getz, G.,Miska, E.A., Alvarez-Saavedra, E., Lamb, J., Peck, D., Sweet-Cordero, A.,Ebert, B.L., Mak, R.H., Ferrando, A.A. et al. (2005) MicroRNA expression profiles classify human cancers. Nature, 435, 834-838.
- Barault, L., Amatu, A., Bleeker, F.E., Moutinho, C., Falcomata, C., Fiano, V., Cassingena, A., Siravegna, G., Milione, M., Cassoni, P. et al. (2015) Digital PCR quantification of MGMT methylation refines prediction of clinical benefit from alkylating agents in glioblastoma and metastatic colorectal cancer. Ann Oncol, 26, 1994-1999.
- Akao, Y., Nakagawa, Y. and Naoe, T. (2006) let-7 microRNA functions as a potential growth suppressor in human colon cancer cells. Biol Pharm Bull, 29, 903-906.
- Preusse, M., Theis,F.J. and Mueller, N.S. (2016) miTALOS v2: Analyzing Tissue Specific microRNA Function. PLoS One, 11, e0151771.
- Sur, I. and Taipale, J. (2016) The role of enhancers in cancer. Nat Rev Cancer, 16,483-493.
https://mp.weixin.qq.com/s/7LPF8P6BZuxYpy6t16drYg

若有收获,就点个赞吧
0 人点赞
