太长不看系列
作者描述和分析了来自 12 只小鼠的八个不同胚胎发育阶段的各组织和器官的甲基化图谱。通过整合 DNA 甲基化与组蛋白修饰、染色质可及性数据,注释了 1808810 个可变甲基化元素,包括大约四分之一的小鼠基因组,并且预测了 468141 个胚胎增强子,这些增强子对应人类基因组各个发育过程中疾病相关的遗传风险基因。这一结果与基因编辑技术相结合可能可以为人类疾病建立新的小鼠模型。胚胎发育的早期阶段,所有组织都出现甲基化减少,而小鼠出生后又出现甲基化显著增加。尽管存在组织异质性,但这种动态变化揭示了一种合理的调控原则,即去除亚稳态抑制性 mCG,可以使基因调控更加快速、灵活。另外,本文延伸了非 CG 甲基化(mCH)的知识,随着 mCH 在胎儿发育后期的进一步积累,位于 mCH 域的基因表达下调。尽管 mCH 的作用仍然不清楚,但现有研究表明,mCH 会直接增加 MeCP2 的结合亲和力。
作者已经做了大量的研究,但目前仍有一些局限性:首先,对于骨骼、生殖腺、胰腺等组织没有研究;其次,本文检测的组织都是异质性的,单细胞表观组的检测可能得到更准确的结果。
延伸阅读
原文链接:https://www.nature.com/articles/s41586-020-2119-x
He, Y., Hariharan, M., Gorkin, D.U. et al. Spatiotemporal DNA methylome dynamics of the developing mouse fetus. Nature 583, 752–759 (2020). https://doi.org/10.1038/s41586-020-2119-x
绘图代码地址1:https://github.com/yupenghe/encode_dna_dynamics
软件代码地址2:methylpy软件:https://github.com/yupenghe/methylpy(流程化工具,好像还挺多人提问的)
Welcome to the home page of methylpy, a pyhton-based analysis pipeline for
- (single-cell) (whole-genome) bisulfite sequencing data
- (single-cell) NOMe-seq data
- differential methylation analysis
软件代码地址3:https://github.com/yupenghe/REPTILE
Regulatory Element Prediction based on TIssue-specific Local Epigenetic marks(好像还没发表)
慢慢看系列
大家好!给大家分享一篇近期发表在 Nature 上的文章,题目是 “Spatiotemporal DNA methylome dynamics of the developing mouse fetus”。文章利用全基因组亚硫酸盐测序(WGBS)分析了来自 12 个小鼠组织或器官的的 9 个不同发育时期(从胚胎形成到成熟)的 168 个甲基化组,通过比较分析,识别出了有 CG 甲基化差异的 1808810 个基因区域,预测了 461,141 个可能的组织特异性增强子,进行了详尽的胚胎发育过程中甲基化时空动态变化的分析。文章的通讯作者为美国索尔克研究所的 Joseph R. Ecker 教授。其主要研究方向为利用基因组和甲基化组测序方法,探索植物、小鼠及人类发育和疾病发生发展中的表观遗传调控。课题组网站为https://ecker.salk.edu/
【研究背景】
哺乳动物胚胎发育涉及精密的基因空间调控。这一过程主要通过复杂的转录结合因子(TFs:transcription factors)与调控 DNA 元素(主要是增强子和启动子)的结合以及表观修饰对这一过程的影响的相互配合来介导。组蛋白及 DNA 的其他修饰会影响转录结合因子接近 DNA。胞嘧啶 DNA 甲基化是一种对基因调控具有重要影响的表观修饰。在哺乳动物中,这一修饰主要发生在 CpG 位点,并且在不同组织和细胞类型的调控元素中是动态的。CpG 位点的胞嘧啶甲基化(mCG)直接影响 DNA 对不同种 TFs 结合的亲和力,并且启动子区域的 mCG 的增加或减少分别与基因表达的增多和减少相关。非 CG 甲基化(mCH,发生在 CpH 位点的胞嘧啶甲基化,H 可以是 A、T、C)也在多种细胞中存在,人神经元中的 mCH 水平甚至超过 mCG。尽管 mCH 的准确功能尚不清楚,但 mCH 可以直接影响 DNA 与 MeCP2(甲基结合蛋白,其突变与雷特综合征相关)的结合。因此,对多种甲基化状态的综合分析具有重要研究意义。
【研究内容】
胞嘧啶 DNA 甲基化在哺乳动物发育过程中受到积极调控。然而,与胚胎移植前的胚胎形成过程相比,中后期的表观遗传学数据非常少,而在这一阶段,主管器官系统的解剖学特征开始出现,人的先天缺陷也变得明显,因而对这一阶段的探索至关重要。作者首先利用全基因组亚硫酸盐测序(WGBS)产生单碱基分辨率的甲基化图谱,并结合已有的转录组图谱,染色质可及性组学数据等进行胚胎发育过程中的动态基因调控分析。
【数据部分】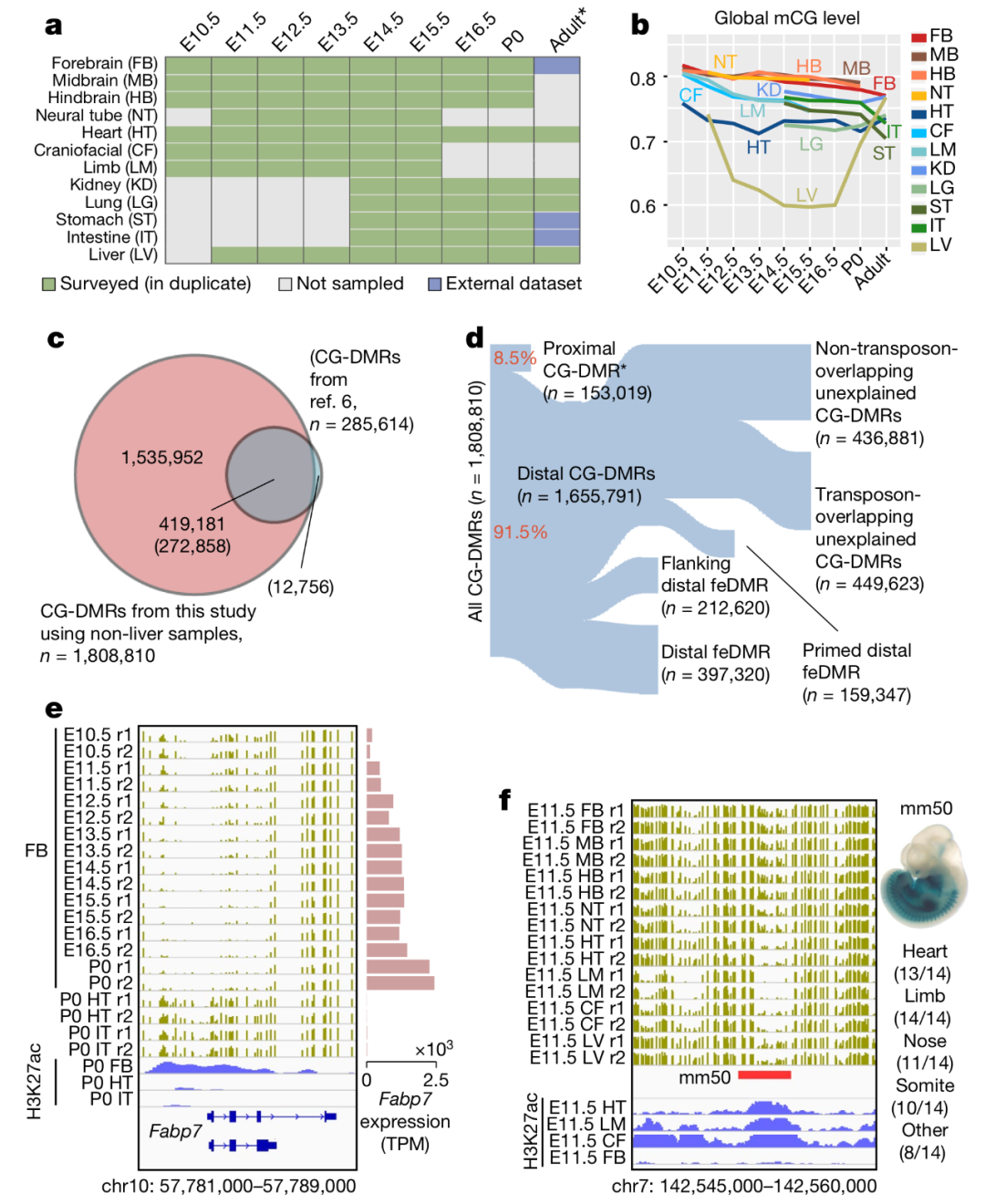
图 1. 小鼠胚胎发育过程中可变甲基化调控元素注释
为了评估发育过程中的小鼠胚胎的胞嘧啶 DNA 甲基化,作者分析了来自大多数主要器官系统和组织类型的 168 个甲基化组(测序深度平均为 31.8x),如图 1a。
所有的甲基化组都呈现高转化率(超过 99.5%),高度可重现性,生物学重复之间的 mCG 定量结果的皮尔森相关系数达到 0.8。其中肝脏甲基化组的相关系数相对较低,这是因为肝脏显示基因组范围内的低甲基化,导致更高的样本差异。如图 1b,胚胎组织的基因组都是高度 CpG 甲基化的,均达到 70-82%,除了肝脏细胞只有 60-74%。
尽管胚胎组织在不同阶段的 mCG 水平差异不大,作者依旧识别出了 1,808,810 个 CG 差异甲基化区域(CG-DMRs),平均 339 bp 长,覆盖了 22.5% 的小鼠基因组。这个综合的胚胎组织 CG-DMR 注释捕获了大约 96% 的之前报道的成年小鼠组织的 CG-DMRs,并且包括大约 150 万个新的区域,如图 1c。
另外,76% 的 CG-DMRs 距离相邻的转录起始位点(TSS)超过 10 kb 远,只有 8.5% 的 CG-DMRs 和启动子、CpG 岛(CGI)、CGI 岸有重叠,这一部分称为近端 CG-DMRs。约 91.5% 的 CG-DMRs 位于远端,并且表现出高度的进化保护,表明他们是功能化的。Fabp7 是一种对于在发育中的大脑中建立桡神经胶质纤维至关重要的基因。实验发现 Fabp7 基因在前脑成熟过程中出现持续的去甲基化,与前脑特异性的乙酰化 H3K27ac 相关。在不同的区域,实验证实 E11.5 的心脏、四肢、鼻子等组织处的增强子呈现去甲基化,如图 1f。
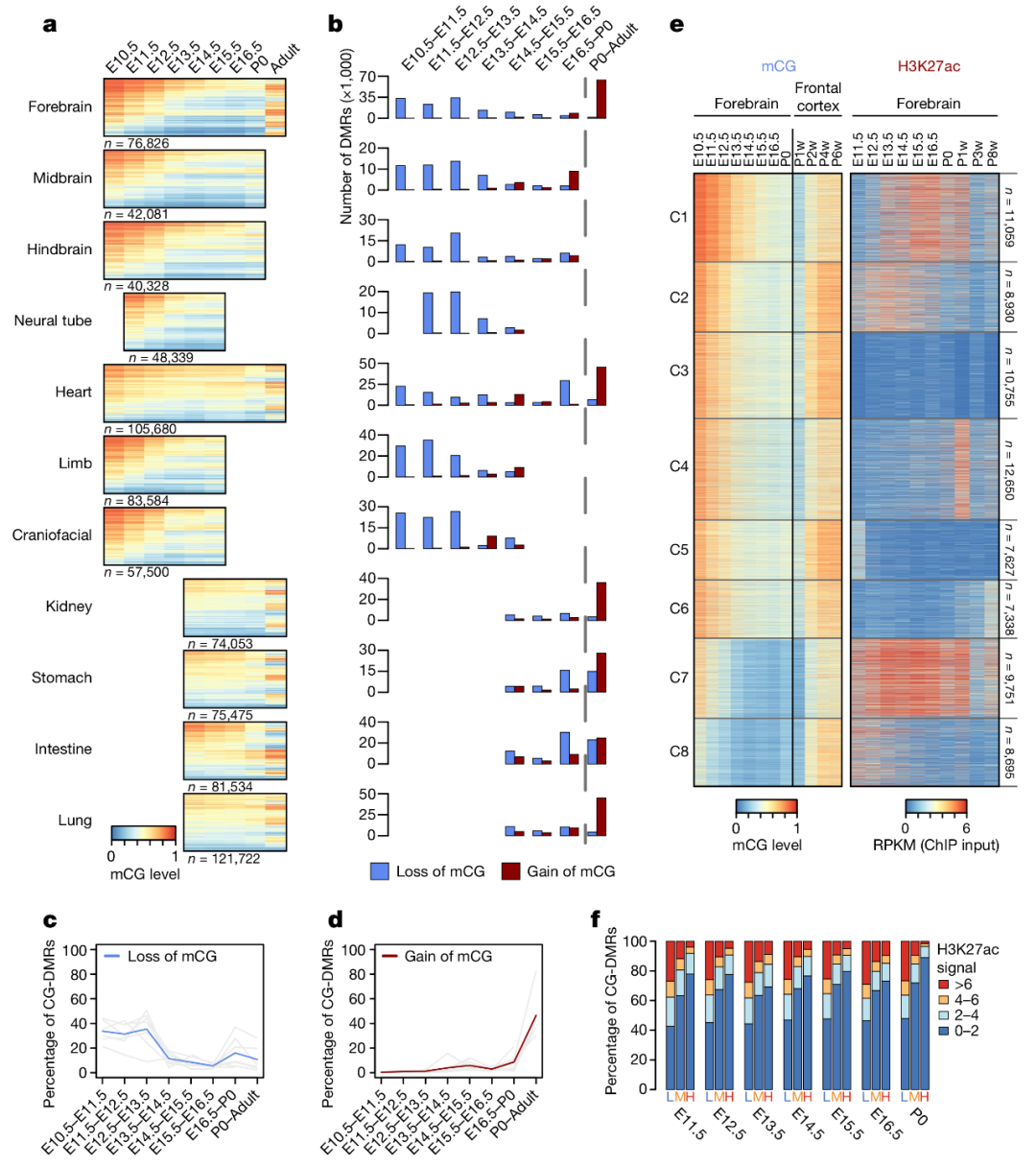
图 2. 组织特异性 CG-DMRs 在胚胎发育中呈现去甲基化,胎儿出生后呈现再甲基化
胚胎发育过程中最主要的甲基化变化是组织特异性的 CG-DMRs 处连续丢失 mCG,覆盖了许多预测的增强子,如图 2a。
然而,这些 CG-DMRs 在胎儿出生后又重新获得 mCG。为了定量每个发育阶段的改变,作者对失去和得到的 mCG 进行了计数,结果如图 2b-d。从 E10.5 到 P0 阶段,77-95% 的 mCG 丢失,其中所有组织(除了心脏)中超过 70% 的丢失发生在 E10.5 到 E13.5。E14.5 处,组织中过半数的细胞的组织特异性 CG-DMRs 均呈现未甲基化。相比 mCG 丢失,胎儿出生后出现 57-86% 的 mCG 获得。
为了进一步指出 CG-DMR 再甲基化的时间及其与增强子活性的关系,作者根据 mCG 和 H3K27ac 活性对前脑组织特异性 CG-DMRs 进行分簇,如图 2e。在所有簇中,mCG 在产后一周到两周显著增加,并且在成年小鼠的组织成熟过程中也进一步增加。作者接下来考察了 mCG 动态变化与预测的增强子活性(根据 H3K27ac 丰度进行评估)之间的关系,尽管 mCG 剥离与 H3K27ac 富集没有必要联系,但高 mCG 指示了低 H3K27ac,图 2e,f。只有 2-9% 的高度甲基化的 CG-DMRs 显示高度 H3K27ac 富集。这些数据表明胚胎发育过程中胞嘧啶甲基化的减少,通过增加 TF 结合或改变组蛋白修饰可能有助于提高增强子活性。
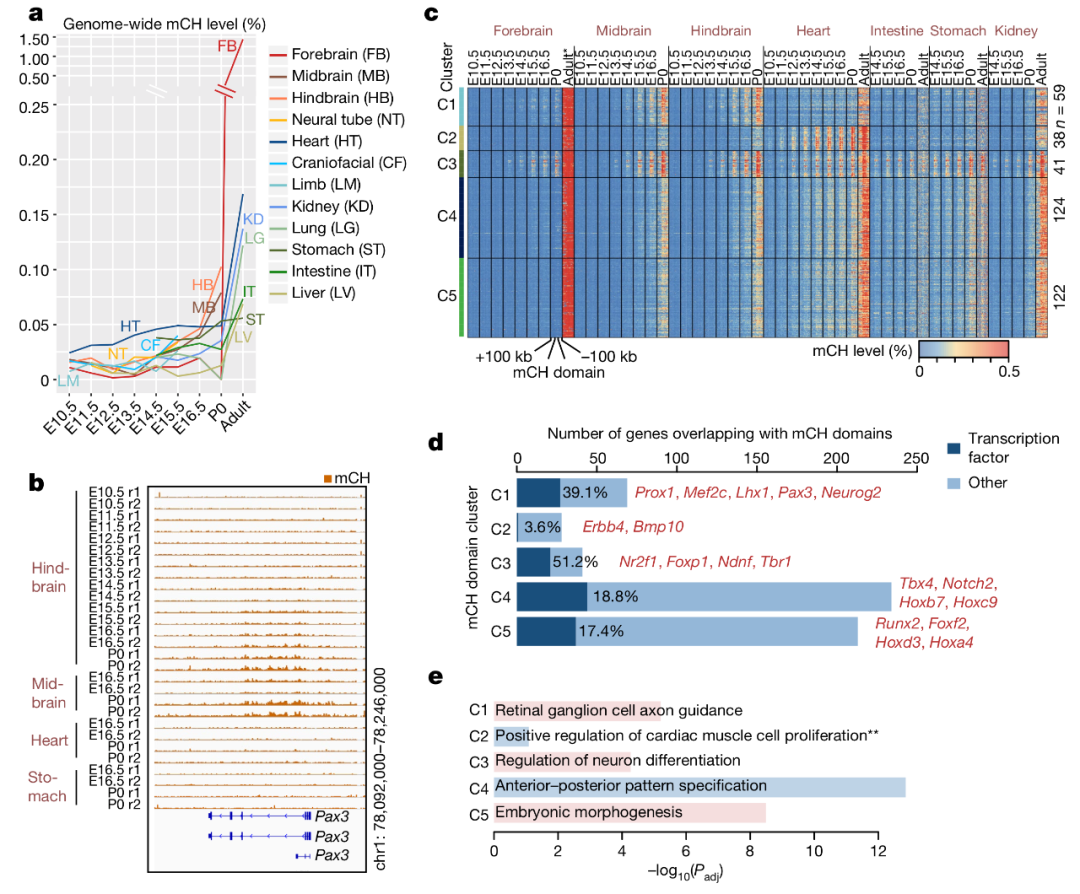
图 3. 基于 mCH 聚集状态预测基因表达水平
目前对于胞嘧啶甲基化中 mCH 的了解依旧知之甚少。本文实验发现在胚胎发育过程中近乎所有组织和器官的 mCH 水平都是可检出的,如图 3a。在脑组织中,mCH 的积累与后脑、中脑、前脑的成熟时间相关。之前的研究已经发现,胚胎干细胞中 mCH 主要是在 DNMT3B 的作用下发生在 5’-CAG-3’的胞嘧啶上,成年组织的 mCH 则主要是在 DNMT3A 的作用下发生在 5’-CAC-3’的胞嘧啶上。在所有的胚胎组织中,mCH 富集在 CAC 位点,并且在组织成熟的过程中特异性增加,表明胚胎具有与成熟组织相似的依赖于 DNMT3A 的 mCH 通路。mCH 聚集优先发生在较大的基因区域,我们称这一区域为 mCH 域,其特点是拥有相比两侧更高的 mCH 水平,如图 3b。作者识别了 384 个 mCH 域,平均长度为 255 kb,其中 92% 的 mCH 域的 61% 的碱基是内含子。22% 的 mCH 域的基因编码 TFs,许多都与组织发育和器官形成有关。为了进一步探索 mCH 聚集的动力学,作者根据 mCH 在胚胎发育过程的动态变化特征将 mCH 域分成五簇,C1-C5,如图 3b,c。C1,C4,C5 的 mCH 域在所有组织中都获得 mCH。值得注意的是,C1 主要富集与神经元分化相关的基因,C4,C5 则主要覆盖与胚胎发育相关的基因,如图 3d,3e。相比 C1、C4、C5 随着胚胎发育在所有组织中都呈现 mCH 水平上升,C2 仅在心脏中出现了这一现象,C3 则是脑特异性的并且覆盖与轴突相关的基因。由于胚胎发育过程中 mCH 聚集在 mCH 域,mCH 域的基因相比这些区域以外的基因更容易被抑制。本文数据显示 mCH 与早期胚胎发育的沉默通路相关。需要开展更多的实验去解释 mCH 的调控机制以及其在转录调控中的作用。
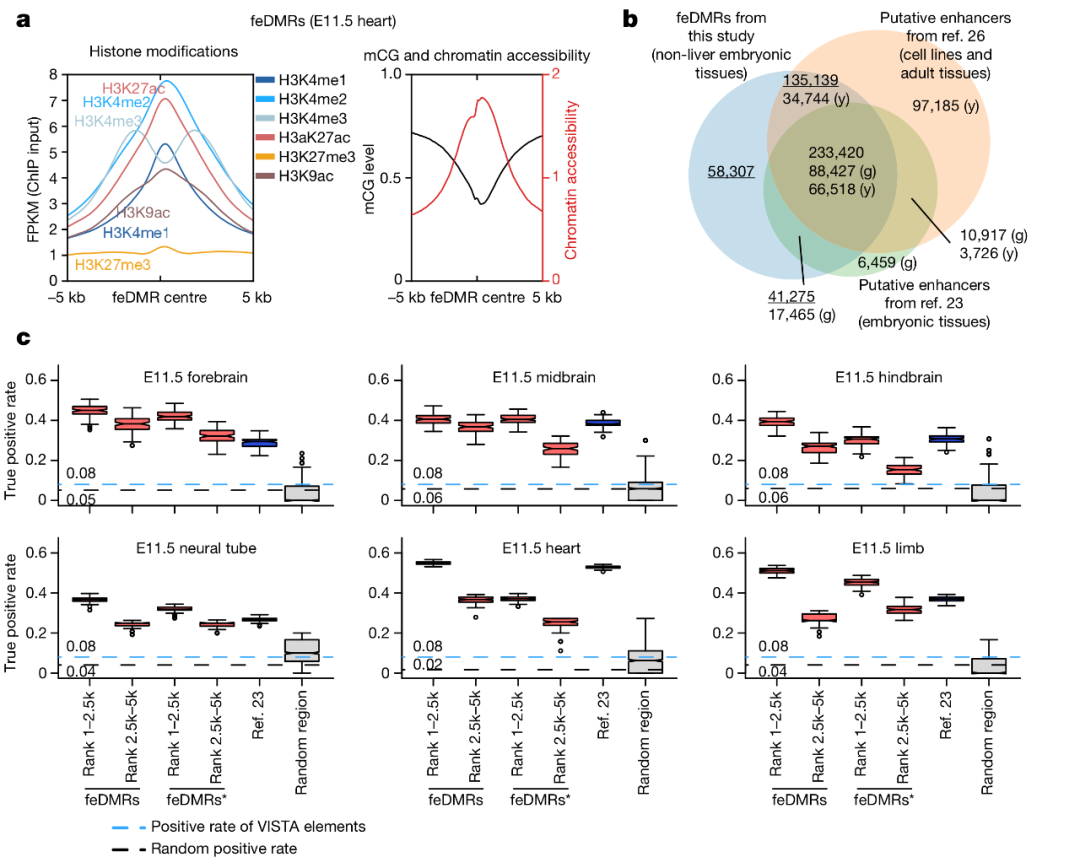
图 4. 发育中的小鼠组织的增强子注释
为了进一步探索胚胎组织发育过程中的动态转录调控,作者利用 REPTILE 算法对 mCG、组蛋白修饰和染色质可及性数据进行了整合,进而对可能与增强子活性相关的胚胎 CG-DMRs 进行了预测。作者识别了 468,141 个候选的 feDMRs(胚胎增强子相关差异甲基化区域)。如图 4a,feDMRs 展示出了类似增强子的染色质特征,包括开放染色质、剥离 mCG,H3K27me3,以及 H3K4me1 和 H3K27ac 富集。其中 21.3% 的 feDMRs 是之前在成熟小鼠组织中没有识别出来的,如图 4b。之后作者进一步证实了其中 37-55% 的 feDMRs 是具有增强子活性的。这些结果与先前的发现一致,证明整合甲基化数据有助于增强子的预测。

若有收获,就点个赞吧
0 人点赞
