哮喘的一个标志性特征是气道中炎症细胞的存在和激活,特别是嗜酸性粒细胞、嗜碱性粒细胞、肥大细胞和 T 淋巴细胞,以及包括气道上皮、成纤维细胞和气道平滑肌在内的结构/驻留细胞的刺激。大量哮喘介质的释放可能会单独或与其他介质协同作用以刺激多种细胞类型,这凸显了气道炎症的复杂性。在对抗原的哮喘急性发作期间,从 IgE 承载细胞释放的预先形成的和脂质介质具有刺激气道平滑肌收缩、神经反射、刺激粘液分泌和支气管壁水肿的潜力,从而导致这种临床表现。即咳嗽、喘息、胸闷和呼气峰流量变化。类似地,抗原呈递给 Th2 记忆细胞会导致细胞因子的产生,这些细胞因子参与气道中炎症细胞的募集、激活和维持,并刺激结构细胞释放生长因子和趋化因子,例如上皮。从各种炎症或结构细胞释放的生长因子以自分泌或旁分泌方式作用,刺激结构细胞增殖,导致杯状细胞增生、气道平滑肌增厚、成纤维细胞增殖和活化、基底膜下胶原沉积和改变的神经反应;统称为气道重塑。因此,细胞因子不仅可以协调炎症细胞向气道的募集,还可以改变气道的结构完整性。
哮喘的一个重要特征是支气管高反应性,指的是哮喘受试者总是对各种刺激更敏感的事实,包括过敏原、污染物、冷空气、运动、蒸馏水和包括缓激肽在内的非生理挑战(组胺和乙酰甲胆碱)。人们普遍认为,气道结构的改变可以解释哮喘患者和健康个体之间气道反应性的差异。在轻度哮喘受试者暴露于空气中的过敏原后,这种基线高反应性可以通过实验 或自然 改变,并且这些事件可以在从环境中去除抗原或使用糖皮质激素治疗后自发逆转。虽然结构重构是基线气道反应性的一个重要决定因素,但轻度哮喘患者对痉挛原的反应性差异很大 以及对不同起始刺激的高反应性表达的差异性,表明除了结构重塑本身之外的机制,有助于基线高反应性。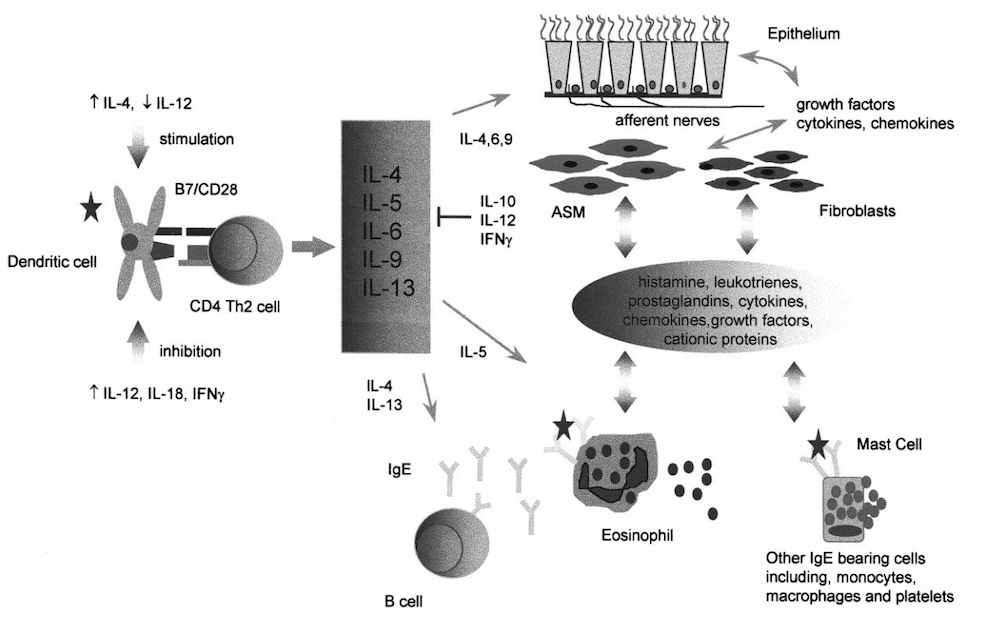
在过去的十年中,人们的兴趣集中在细胞因子作为重要的哮喘介质的作用,以及越来越多的人认识到免疫系统在协调潜在的临床病理学中的作用。 本综述将重点关注各种细胞因子在气道炎症中的作用,特别是采用鼠模型的研究。
T淋巴细胞在过敏性炎症中的作用
不同的 T 淋巴细胞功能(细胞溶解、细胞凋亡的诱导、B 细胞帮助和炎症细胞募集)由不同的 T 淋巴细胞亚群介导,这些亚群可以通过它们分泌的受体分子(细胞因子)来识别。 CD4+ T 淋巴细胞被归类为 T 辅助 1 (Th1) 细胞,可提供对病原体(如结核分枝杆菌)的免疫力;作为 T 辅助 2 (Th2) 细胞,会引起过敏性炎症。 Th1淋巴细胞分泌IFNg、IL-2和TNF-b,而Th2淋巴细胞分泌细胞因子,如IL-4、IL-5、IL-6、IL-9、IL -10 和 IL-13。包括 IL-4 和 IL-13 在内的细胞因子涉及 B 细胞的同种型转换以产生 IgE ,IL-5 促进嗜酸性粒细胞的分化、募集和存活 ,因此被认为在气道炎症中起重要作用。一个亚群产生的细胞因子能够拮抗另一个亚群的生长和功能,IFN-g 和 IL-4 分别抑制 Th2 和 Th1 淋巴细胞的生长和功能。
在哮喘患者的气道黏膜下层和痰液中,以及在过敏原诱导的晚期哮喘反应中,表达 IL-4 和 IL-5 mRNA 但不表达 IFN-g 的 CD4+ T 淋巴细胞数量增加 嗜酸性粒细胞数量增加,尽管在一些研究中,在支气管肺泡灌洗液 (BAL) 中恢复的 T 淋巴细胞的细胞内染色证明了 IFN-g 分泌细胞占优势。 因此,细胞因子在哮喘发病机制中的确切作用仍有待确定,然而,小鼠模型的使用使研究人员能够使用包括单克隆抗体和靶向基因破坏在内的多种技术来探索炎症反应的复杂性。
在先前用针对 CD4 的单克隆抗体治疗的小鼠中,抗原攻击后 24-48 小时的肺嗜酸性粒细胞募集显著受损。 有趣的是,抗原攻击后 9 小时嗜酸性粒细胞向肺的募集没有受到抑制,这表明 CD4+ T 淋巴细胞独立机制,例如 肥大细胞可以在早期将嗜酸性粒细胞募集到气道中发挥作用。 在 CD4+ T 淋巴细胞耗尽后,免疫小鼠的抗原攻击后增加的气道对乙酰甲胆碱的反应也被消除。 总之,这些研究表明 T 淋巴细胞是协调气道炎症小鼠模型中组织嗜酸性粒细胞增多和支气管高反应性的重要细胞。
最近,研究人员使用过继转移技术进一步探索 T 淋巴细胞在过敏性炎症中的作用。 TCR-ab 特异性 Th0 和 Th2 克隆过继转移到 SCID 或 BALB/c 小鼠中,在抗原攻击后诱导肺嗜酸性粒细胞增多和支气管高反应性。 相比之下,卵白蛋白特异性 Th1 细胞克隆过继转移到小鼠体内会导致中性粒细胞和单核细胞渗入肺中,而不会引起支气管高反应性。 类似地,遗传性高反应性小鼠的 T 淋巴细胞过继转移可以增加低反应性受体小鼠的气道反应性,尽管导致这些结果的 T 淋巴细胞因子仍有待确定。 这些研究证明了抗原特异性 Th2 淋巴细胞在协调嗜酸性粒细胞募集和促进支气管高反应性中的关键作用。
T 淋巴细胞活化的一个关键步骤是通过抗原呈递细胞(包括树突细胞、巨噬细胞和 B 细胞)呈递外来抗原。在 MHC II 分子上以肽片段的形式呈递加工过的抗原,以及包括 B7-1 (CD80) 和 B7-2 (CD86) 在内的共刺激配体的表达,为 T 淋巴细胞识别、激活提供了必要的信号和克隆扩增[29]。树突状细胞在 T 淋巴细胞的激活中起主要作用,这一事实证明,肺树突状细胞在刺激幼稚抗原特异性 T 淋巴细胞方面远优于 B 细胞 [30],而 B 细胞和巨噬细胞可以将抗原呈递给以前在体外激活的 T 淋巴细胞 [31],许多体内研究表明,这些细胞在气道炎症中的抗原呈递中起次要作用。因此,支气管高反应性和肺嗜酸性粒细胞增多可在没有 B 淋巴细胞的情况下发生,而巨噬细胞通过抑制树突状细胞来下调 T 细胞功能 [35]。相比之下,更昔洛韦治疗在髓系树突细胞谱系细胞中表达单纯疱疹病毒 I 胸苷激酶 (TK) 基因的小鼠后,有条件地消融树突细胞,抑制致敏小鼠抗原攻击后嗜酸性粒细胞向肺的募集。因此,嗜酸性粒细胞的募集依赖于树突状细胞向记忆 T 淋巴细胞的抗原呈递。与这些研究一致的是,较早的报告显示用可溶性融合蛋白 (CTLA4-Ig) 进行治疗,该蛋白在致敏小鼠的抗原攻击之前识别共刺激分子 CD80 和 CD86;最近在 CD28 基因敲除小鼠中,表现出对吸入抗原的免疫反应减弱,表现为 Th2 细胞因子产生的显着抑制、支气管高反应性和伴随的嗜酸性粒细胞增多。同样,针对 CD86的单克隆抗体在免疫小鼠的抗原攻击后抑制 Th2 细胞因子的产生、支气管高反应性和随之而来的肺嗜酸性粒细胞增多。然而,同样清楚的是,针对 CD80 的抗体会部分减弱嗜酸性粒细胞向气道的募集,尽管不影响 Th2 细胞因子的产生。因此,这种治疗对嗜酸性粒细胞募集的影响可能是这种抗体直接作用于已知表达 CD80 的嗜酸性粒细胞的结果。或者,这些分子在不同细胞上的时空表达也可能解释了文献中观察到的一些不一致。同样,虽然一些研究报告称,在抗原攻击前给予抗 CD86 仅部分有效地减轻支气管高反应性和嗜酸性粒细胞的募集,但在其他研究中并非如此,并且这些差异可能与用于致敏小鼠的抗原和佐剂剂量以及攻击方案中使用的抗原剂量有关。鉴于新的共刺激 B7 分子和诱导型共刺激物 (ICOS) 的发现,需要进一步研究其他共刺激分子在抗原呈递中的潜在作用。然而,普遍的共识是,通过树突状细胞上的 CD28/B7 分子发出的信号是小鼠炎症模型中 Th2 细胞因子产生、IgE 产生、嗜酸性粒细胞募集和支气管高反应性的重要决定因素。
IL-4和IL-13
IL-4 已被证明对 Th2 淋巴细胞定向分化和 B 细胞向 IgE 表型的同种型转换很重要,尽管 IL-4 在过敏原诱导的嗜酸性粒细胞募集和支气管高反应性中的作用是有争议的。 因此,在 IL-4 基因破坏的小鼠或在抗原免疫期间用 IL-4 单克隆抗体治疗的小鼠中,嗜酸性粒细胞募集显着减弱,从而支持 IL-4 是一种重要的 Th2 淋巴细胞定型的决定因素。 在此过程中 IL-4 的可能来源一直是备受争议的主题,研究调查了 gd T 淋巴细胞、肥大细胞 和 NK1.1+ T 淋巴细胞的作用。 其他研究表明,在致敏小鼠的抗原攻击过程中产生的 IL-4 对于将嗜酸性粒细胞募集到气道很重要,因为这种反应可以在事先给予 IL-4和 IL-4 受体的单克隆抗体后被抑制。
相比之下,其他研究表明,在抗原致敏或抗原攻击之前使用针对 IL-4 的抗体进行治疗对嗜酸性粒细胞向气道的募集没有影响。有趣的是,最近的一项研究表明,在 IL-4 缺陷小鼠或从 IL-4 缺陷小鼠过继转移 Th2 细胞克隆后的小鼠中,嗜酸性粒细胞仍会募集到肺部。然而,在这两种情况下,嗜酸性粒细胞从组织迁移到管腔似乎受到了损害,尽管这种现象的机制仍有待确定。同样,IL-4 依赖性和独立机制都有助于在小鼠炎症模型中观察到支气管高反应性。这些差异不能仅仅通过使用不同的小鼠品系来解释,因为使用具有相似遗传背景的小鼠已经出现了许多相互矛盾的数据。一个更合理的解释是,在所采用的不同实验条件下,CD4+ T 淋巴细胞可能通过 IL-4 依赖和独立途径诱导支气管高反应性和嗜酸性粒细胞增多。这一结论进一步得到证实,Th2 淋巴细胞亚群的发育和定型不依赖于 IL-4。
许多研究进一步研究了阻断 IL-4 受体信号传导的作用,试图进一步阐明 IL-4 在过敏性炎症中的作用。 IL-4 激活多种信号通路,包括信号转导和转录激活因子 (STAT),它们属于被 Janus 激酶磷酸化的酪氨酸蛋白家族。 缺乏 STAT6 的小鼠无法引发显着水平的卵清蛋白特异性 IgE 或 IgG1、Th2 细胞因子(包括 IL-4 和 IL-5)水平升高、肺嗜酸性粒细胞募集或抗原攻击后支气管高反应性 ,这表明在 STAT6 缺陷小鼠中 Th2 细胞的定型已受到损害。
粘液分泌过多是哮喘的一个特征,可能导致气道受限,许多研究也在气道炎症的小鼠模型中记录了这一特征。来自免疫的野生型小鼠的上皮细胞在抗原攻击后对粘蛋白染色呈阳性,通过阻断 IL-4 受体和 IL-4 转基因小鼠表现出继发于粘液糖蛋白含量增加的上皮细胞肥大。然而,在将 Th2 克隆从缺乏 IL-4 的小鼠过继转移到野生型后,也可以在小鼠中引发粘蛋白产生,但在缺乏 IL-4 受体 (IL-4Ra) α 链的小鼠中则不会。 ),它也参与 IL-13 信号传导,因此暗示 IL-13 在粘蛋白产生中的作用。这些研究突出了这些模型中不同细胞因子所起的补偿作用。还研究了 STAT6 在这种反应中的作用,似乎在 STAT6 缺乏的小鼠中,上皮细胞产生的粘蛋白被消除了。虽然粘蛋白的产生可能对基线肺阻力产生重要影响,但它似乎在气道高反应性中没有发挥重要作用。此外,在抗原攻击前给予 IL-5 后,在 STAT6 缺乏的小鼠中,支气管高反应性和肺嗜酸性粒细胞增多症可以重建。这些数据表明粘蛋白的产生对于这些模型中的支气管高反应性不是强制性的,但显然对于哮喘恶化期间的气流受限很重要。需要进一步的研究来确定抑制 STAT6 是否具有临床用途,特别是如果 IL-4 非依赖性 CD4 T 细胞和其他炎症细胞提供 IL-5 的来源。事实上,在缺乏 STAT6 的 BALB/c 小鼠中,嗜酸性粒细胞向肺的募集仅被抑制了 50%,这表明 Th2 承诺不仅仅依赖于 IL-4,并且与显示 Th2 反应发展的研究一致在 BALB/c 小鼠中可以独立于 IL-4发生。
虽然很明显 IL-4 信号传导可以在 STAT6 缺陷小鼠中被破坏,但通过该途径发出信号的其他细胞因子的作用可能解释了在检查 IL4 破坏对过敏的影响的研究中观察到的一些差异。现在认识到,IL-13 信号转导是通过一个复合物发生的,该复合物涉及将 IL-4R 的 a 链募集到 IL-13Ral 和 STAT6 依赖性途径。因此,IL-13 可以与 IL-4 具有重叠的生物学特征 [86]。 IL-13 是一种多营养细胞因子,可诱导人类 B 细胞经历 lg 同种型转换为 IgE [11];促进 Th2 淋巴细胞分化 [87],抑制 T 淋巴细胞产生 IFNg [86];增加小鼠巨噬细胞 [88] 和人气道上皮 [89] 中脂氧合酶的表达;并增强体外人嗜酸性粒细胞的活化[90]。最近的研究表明,与缺乏 IL-4 的小鼠相比,IL-4Ra 对免疫反应的抑制程度明显更大,并表明通过该受体的 IL-13 信号传导在气道炎症中起作用。 91]。最近,已经表明针对 IL-13 的抗体在免疫小鼠的抗原攻击后抑制粘蛋白产生和气道高反应性 [92,93],尽管对嗜酸性粒细胞募集的影响存在争议。相反,在未免疫小鼠中,在没有抗原攻击的情况下,气道中 IL-13 的过表达导致粘液分泌过多、嗜酸性粒细胞分泌、嗜酸性粒细胞募集和气道高反应性 [94]。同样,IL-13 的外源性给药通过依赖 IL-4Ra 的途径刺激粘蛋白产生 [92];上调 IgE 的产生 [93];并诱导嗜酸性粒细胞募集 [92,93]。这些研究共同表明 IL-4 和 IL-13 在过敏性炎症过程中具有补偿作用,并且必须去除这两种细胞因子才能完全下调炎症反应。然而,很明显,IL-4 和 IL-13 的作用更复杂,这从同时破坏两种细胞因子的研究中可以看出。在寄生虫感染 (N. brasiliensis) 的动物中,双敲除菌株的肉芽肿形成、嗜酸性粒细胞募集、Ig 产生和 Th2 细胞因子产生比单独破坏任一基因的程度要大得多,这表明这些细胞因子在在寄生虫感染中产生 Th2 反应 [95]。尽管如此,尽管 Th2 反应显着减弱,但与野生型小鼠相比,IL-4/IL-13 缺陷小鼠的胃肠道蠕虫排出仍然很明显,尽管寄生虫排出延迟。有趣的是,IL5 产生和嗜酸性粒细胞募集在这些敲除小鼠中仍然很明显,这表明 IL-4/IL-13 独立机制也可以在蠕虫排出和嗜酸性粒细胞募集中起作用[95]。鉴于发现 Th2 反应仍然可以,独立于 IL-4 和 IL-13 [96] 引发。
IL-5
鉴于其在嗜酸性粒细胞的分化募集和存活中的重要作用,另一个受到相当大关注的细胞因子是 IL-5 [12±14]。 大量研究强调了 IL5 在小鼠气道炎症模型中的重要性。 在抗原攻击之前施用针对 IL-5 的单克隆抗体可减轻过敏小鼠的支气管高反应性和随之而来的嗜酸性粒细胞募集 [20,72,98,99]。 此外,抗原攻击后的支气管高反应性和肺嗜酸性粒细胞增多在免疫抗原的 IL-5 基因破坏小鼠中被消除 [14],但可以在过继转移分泌 IL-5 的 CD4 T 淋巴细胞后重建 [100]。 此外,如果 IL-5 在非致敏小鼠的肺中过度表达,则会观察到气道嗜酸性粒细胞增多和支气管高反应性 [101]。
虽然有压倒性的证据支持 IL-5 在将嗜酸性粒细胞募集到气道和支气管高反应性中的关键作用,但许多模型表明,除 IL-5 之外的因素可以在这些反应中发挥重叠作用。因此,针对 IL-5 的单克隆抗体在减少对气道的嗜酸性粒细胞募集的同时,在抗原激发之前 [79] 或在确定的气道嗜酸性粒细胞增多 [102] 之后施用时未能减轻支气管高反应性。类似地,虽然之前已经证明 IL-5 缺乏的 C57B1/6 小鼠在抗原攻击后不会出现肺嗜酸性粒细胞增多和支气管高反应性 [14]。 IL5 敲除 BALB/c 小鼠还记录到气道嗜酸性粒细胞显着减少并且气道没有任何显着的形态学变化,但是,支气管高反应性仍然很明显 [23]。同样,虽然缺乏 IL-5 的小鼠在初次感染期间排出寄生虫的能力较低,但缺乏 IL-5 并不会影响这些小鼠在随后的暴露中清除寄生虫的能力 [103]。在 IL-5 敲除小鼠的炎症部位发现的嗜酸性粒细胞的残留水平是否仍然足以在这些模型中表达支气管高反应性 [23] 或驱除寄生虫 [103] 仍有待确定。
因此,很明显,某些小鼠模型中的支气管高反应性可以独立于 IL-5 引发,然而,T 淋巴细胞似乎对这种反应至关重要,因为针对该细胞的单克隆抗体消除了这种反应 [23]。 这些 T 细胞衍生因子的性质仍有待确定,但包括 IL-13 [92,93] 和 IL-9 在内的候选因子(见第 6.2 节)仍有待在这些模型中进行研究,很明显,各种小鼠的遗传背景 在没有 Ig 和 IL-5 依赖途径的情况下,菌株是气道高反应性的重要决定因素 [71]。
嗜酸性粒细胞活化状态的差异也可能是这些研究中报告的相互矛盾的结果的重要决定因素。有趣的是,尽管存在支气管高反应性 [98,101,104,105],但募集到肺部的嗜酸性粒细胞似乎并未证明气道中有细胞外颗粒产物沉积的证据,尽管这不是一个普遍的发现 [23,72]。虽然许多研究表明 IgE 和 IL-5 有助于小鼠气道高反应性的发展 [33,70,98],但这不太可能涉及 IgE 通过嗜酸性粒细胞上的抗原交联,因为鼠嗜酸性粒细胞似乎缺乏高和低位 IgE 受体,但确实表达 IgG 受体 [56,106]。因此,很明显,不同的小鼠模型可以证明 IL-5 依赖和独立机制导致气道炎症,这一发现在寄生虫感染的小鼠模型中得到了很好的证实 [103,107] 并突出了复杂的性质导致支气管高反应性发展的机制,并且是确定哪些模型密切模仿人类哮喘的挑战。哮喘患者吸入人重组 IL-5 与气道对乙酰甲胆碱的敏感性增加三倍有关。攻击后 24±48 小时,痰嗜酸性粒细胞数量增加 6 倍,痰嗜酸性粒细胞阳离子蛋白水平增加 3 倍 [108]。相比之下,针对 IL-5 的单克隆抗体虽然显着减少了痰液和血液中的嗜酸性粒细胞数量,但未能改变哮喘患者的晚期哮喘反应或对乙酰甲胆碱的支气管高反应性 [109]。这项研究表明,IL-5 独立途径可能与哮喘的晚期哮喘反应和基线支气管高反应性有关。其他针对 IL5 的单克隆抗体是否证明更有效、慢性治疗是否证明更有效和/或这种治疗与常规抗炎药联合是否在临床环境中占有一席之地仍有待确定。显然,需要进一步的研究来阐明 IL-5 在哮喘中的确切作用。
IL-6、IL-9、IL-10
其他 Th2 细胞因子的作用也已在小鼠炎症模型中进行了研究。
气道上皮细胞中 IL-6 的过度表达与淋巴细胞渗入气道有关,然而,观察到气道对乙酰甲胆碱的敏感性降低,这表明 IL-6 可能在气道生理学中具有保护作用 [110]。 IL-6 在过敏原激发后气道炎症中的作用仍有待确定。
鉴于 IL-9 基因在人类 5 号染色体(小鼠,13 号染色体)上的位置以及染色体 5q31-q33 在特应性疾病中可能存在的联系 [111] 导致研究人员研究这种细胞因子在小鼠模型中的作用的信息。似乎小鼠支气管对乙酰甲胆碱反应的遗传差异可能是 IL-9 表达水平不同的结果[111],事实上,IL-9 在气道中的过度表达与显着相关。此外,IL-9 转基因小鼠气道的特征是淋巴细胞、嗜酸性粒细胞和肥大细胞数量增加,以及气道重塑的证据,包括上皮下胶原沉积 [112]。此外,通过更多的嗜酸性粒细胞募集来评估,气道内 IL-9 的过度表达似乎会加剧气道炎症。总 IgE;和非转基因小鼠相比,抗原攻击后的支气管高反应性[113]。特别令人感兴趣的是,即使在没有抗原攻击的情况下,在肺部也发现了大量的嗜酸性粒细胞 [112±114],这可能是 IL-9 刺激趋化因子产生能力的结果,包括来自气道上皮的 eotaxin、MIP-1a 和 MCP1、3 和 5 [114]。
Interleukin-10 由 Th2 细胞、B 细胞、单核细胞和巨噬细胞产生,抑制 Th1 细胞和迟发型超敏反应;巨噬细胞功能;和许多促炎细胞因子和趋化因子的产生,正是这些后一种效应被认为是治疗过敏性疾病的可能治疗靶点[115]。对炎症过敏模型的研究表明,IL-10 与卵白蛋白同时给药,显着减少了免疫小鼠气道中中性粒细胞和嗜酸性粒细胞的募集,继发于抑制 TNFa 的产生 [116]。仅当 GMCSF 基因在肺中过度表达时,对小鼠的反复气溶胶攻击显示支气管高反应性和肺嗜酸性粒细胞增多 [117]。在这种情况下,在这些小鼠的肺中施用携带 IL10 转基因的构建体显着减弱了 BAL 液中卵白蛋白诱导的支气管高反应性、肺嗜酸性粒细胞增多和细胞因子含量(IL-4、IL-5 和 TNFa)。最后,靶向破坏 IL-10 基因,增加了淋巴细胞和嗜酸性粒细胞向气道的渗透,以及在鼻内暴露于烟曲霉后 IL-5 的释放 [118]。这些研究共同支持这样的观点,即 IL-10 可以在许多小鼠炎症模型中作为过敏反应的内源性抑制因子。
IL-12、IL-18、IFN-γ
最近的研究调查了 Th1 和 Th2 细胞克隆的过继转移的影响,以试图抑制小鼠中 Th2 介导的过敏性炎症。 虽然利用急性抗原攻击方案的研究未能证明 Th1 细胞克隆抑制过敏性炎症的能力,事实上,在某些条件下似乎会加剧这种反应 [26,27],但最近表明 Th1 细胞可以减弱 慢性抗原暴露后 Th2 细胞介导的炎症 [119] 并表明增强 Th1 反应可诱导体内免疫偏差。 这一观点与用卡介苗 (BCG) [120±122] 和细菌 DNA 寡脱氧核苷酸基序 (CpG-ODN) [123±125] 治疗后过敏性炎症得到抑制的结果一致。 因此,Th1 细胞因子在调节过敏性炎症中的作用已经在各种小鼠模型中进行了研究,以考虑新型抗感染疗法潜力。
IL-12 是一种由抗原呈递细胞产生的异二聚体细胞因子,其最显着的作用是上调 Th1 免疫反应,从而促进针对细胞内微生物感染的免疫保护 [126],并为下调Th2介导的过敏反应。在抗原攻击前给予 IL-12 可不同程度地抑制气道嗜酸性粒细胞增多、淋巴细胞产生 Th2 细胞因子和支气管高反应性 [127±131]。相比之下,IL12 作为佐剂的施用,即在抗原致敏过程中,试图改变对 Th1 表型的免疫反应,在某些模型中,在过敏原挑战后不会抑制嗜酸性粒细胞向气道的募集 [131],这表明用 IL-12 进行辅助治疗在这方面效果较差。然而,在适当的时间在射出部位控制足够水平的 IL-12 的能力可能会影响免疫时向 Th1 表型的免疫偏差程度 [128,130]。在抗原暴露前 IL-12 基因过表达 [132,133] 或用热灭活李斯特菌疫苗接种后刺激内源性 IL-12 释放后,这些药代动力学和药效学不确定性得以克服 [134]。在这种情况下,观察到抗原攻击后肺嗜酸性粒细胞增多的抑制。与这些研究一致的是,IL-12 缺乏小鼠表现出增强的嗜酸性粒细胞向气道募集,BAL 液中 IL-4、TNFa 和血清 IgE 水平升高[126];在抗原攻击之前施用的针对 IL-12 的抗体增强了炎症反应 [135]。
由于 IL-12 是 IFNg 产生的有效诱导剂,因此已经研究了这种细胞因子在 IL-12 的免疫调节作用中的作用。因此,IL-12 对抗原致敏小鼠肺嗜酸性粒细胞增多的抑制作用在用针对 IFNg 的抗体治疗后减弱 [127,129]。同样,IL-12 减弱嗜酸性粒细胞向气道募集的能力在 IFNg 受体缺陷小鼠中并未完全逆转 [136],这表明 IFNg 依赖和独立机制可以解释 IL12 对过敏反应的抑制作用。信息。在抗原暴露之前 [133] 或在先前致敏小鼠的抗原攻击之前 [132] 的气道中过度表达 IL-12 的研究中发现了这些多种途径的证据。 IL-12 在抗原免疫后但在攻击之前通过痘苗病毒在气道中的过表达抑制了 IL-4 和 IL-5 的局部产生,同时 IFNg 水平增加,并抑制了嗜酸性粒细胞募集和支气管高反应性。在 IFNg 受体缺乏的小鼠中,IL-12 诱导的支气管嗜酸性粒细胞增多和高反应性抑制被逆转,这表明 IFNg 在这种反应中的作用[132]。相比之下,在抗原致敏之前利用腺病毒介导的基因转移技术,在 GMCSF 环境中卵清蛋白诱导的嗜酸性粒细胞增多被很大程度上不依赖于 IFNg 的 IL-12 的过表达显着抑制,而 IL-12 抑制 IgE 的产生。 12 是 IFNg 依赖性的 [133]。最后,在抗原攻击之前外源性给予 IL-12 会增加 IFNg 受体敲除小鼠中卵清蛋白诱导的嗜酸性粒细胞增多 [136]。因此,似乎 IL-12 暴露的时机对于启动涉及 IFNg 的途径至关重要,并且在某些情况下,可能会加剧气道炎症 [136]。在这方面,旨在提高内源性 IL-12 水平的机制可能会提供更好的方法。事实上,低剂量 CpG-ODN 抑制 Th2 反应的能力取决于 IL-12 的内源性释放 [137]。然而,更高剂量的这种物质也独立于 IL-12 和 IFNg 抑制 Th2 反应,这表明在适当的条件下可以引发其他途径来干扰 Th2 反应,例如。 IL-10 和/或抗原呈递细胞上共刺激表达的改变 [137,138]。这可能对针对免疫偏差的策略产生重要影响,作为基于 IL-12 依赖性途径治疗过敏性炎症的治疗方法,并可能解释 IL-12 在哮喘治疗中的失败 [139]。
IL-18 是最近描述的一种细胞因子,它是 IFNg 产生和致敏 T 细胞中 Th1 细胞分化的有效诱导剂 [140],因此可能在调节 Th2 过敏反应中发挥作用。 IL-18 在气道上皮中组成型表达,并且在过敏小鼠的抗原攻击后似乎下调,这可能是 Th2 细胞因子抑制作用的结果 [141]。虽然 IL-18 的全身给药对气道炎症没有影响,但与 IL-12 的共同给药显着减弱了抗原诱导的支气管高反应性、肺嗜酸性粒细胞增多和 Th2 细胞因子的产生 [142]。相比之下,在本研究中,IL-18 与抗原的局部共同给药增加了嗜酸性粒细胞的募集 [143]。有趣的是,针对 IL18 的抗体的全身给药促进了嗜酸性粒细胞在早期而不是晚期的肺部募集,并且与抗原攻击后支气管高反应性的增加有关,这表明 IL-18 产生的系统性抑制嗜酸性粒细胞募集并抑制支气管高反应性[143]。与这一发现相反,IL-18 的全身给药增强,而 IL-12 抑制,嗜酸性粒细胞募集到过敏小鼠的气道和 IL-18 并没有改变该模型中抗原诱导的支气管高反应性 [144]。 IL-18 的促嗜酸性粒细胞活性归因于 INFa [144] 和 eotaxin [143]。显然,需要进一步的工作来阐明 IL-18 在小鼠过敏性炎症中的作用,并再次强调细胞因子生物学的复杂性,它依赖于炎症反应期间这些细胞因子的空间和时间释放。
源自 CD4+、CD8+ T 淋巴细胞和 NK 细胞的 IFNg 在 Th1 介导的免疫中起重要作用,并抑制 Th2 细胞的增殖,从而调节 Th2 介导的反应。 IFNg 的外源性给药抑制了免疫小鼠中 IgE 的产生、嗜酸性粒细胞和 CD4+ T 淋巴细胞的募集以及抗原诱导的支气管高反应性 [145±150],而 IFNg 在气道中的过表达也抑制了过敏性炎症反应 [151]。针对 IFNg 的抗体对 [152] 或增强 [145] 嗜酸性粒细胞向气道的募集无效。这些不一致是否与使用的不同模型或抗体中和 IFNg 的效率有关尚不清楚,因为抗体已显示可减弱抗原诱导的支气管高反应性 [152]。涉及干扰 IFNg 信号传导的研究已被用于进一步阐明这种细胞因子在气道炎症中的作用。缺乏 IFNg 的小鼠在抗原攻击前增加了基线气道反应性,尽管抗原攻击后对气道的嗜酸性粒细胞募集受损,并且由于体外 IFNg 不会损害 Th2 细胞因子的产生,因此得出结论,在某些情况下 IFNg可能促进嗜酸性粒细胞功能 [150],最近的研究支持 IFNg 可以促进嗜酸性粒细胞存活的观点 [153]。与这些发现缺乏 IFNg 受体的小鼠相比,在抗原攻击后保留了募集嗜酸性粒细胞的能力 [136,154],有趣的是,BAL 液中的嗜酸性粒细胞炎症和 Th2 细胞因子水平延长了 [154],这表明IFNg 可以通过抑制肺中 Th2 细胞产生的 Th2 细胞因子或通过减少 CD4 T 淋巴细胞向肺的募集来干扰嗜酸性粒细胞炎症 [145,146]。 Th2 细胞过继转移到过表达 IFNg 基因的小鼠中,其特点是抗原攻击后 BAL 液中的嗜酸性粒细胞和 Th2 细胞因子含量减少,因此暗示了这种细胞因子对嗜酸性粒细胞募集的抑制作用 [151]。同样,在 Th1 和 Th2 细胞共转移后,肺嗜酸性粒细胞增多受到抑制,这种效应取决于 Th1 细胞数量,是 IFNg 释放的结果 [119]。在 Th1 与 Th2 淋巴细胞共转移后,BAL 液体中 IL-4、IL-5 和 IL-13 的水平没有降低,这表明 IFNg 对嗜酸性粒细胞募集的抑制作用也可能发生在 Th2 释放细胞因子的下游细胞[119],尽管机制仍有待建立。主要共识是 IFNg 依赖途径倾向于抑制小鼠的 Th2 过敏反应,然而,在某些情况下,此类途径可能促进这些反应的可能性需要澄清 [150,152]。
信源:Riffo-Vasquez Y, Pitchford S and Spina D 2000 Cytokines in airway inflammation Int J Biochem Cell Biol 32 833–53

若有收获,就点个赞吧
0 人点赞
